Fungal Endophytes: Drought Tolerance in Plants: Difference between revisions
SBarnes7151 (talk | contribs) No edit summary |
SBarnes7151 (talk | contribs) No edit summary |
||
| Line 1: | Line 1: | ||
{{Uncurated}} | {{Uncurated}} | ||
<br>It has been estimated that over 80% of terrestrial plants form a symbiotic association with fungi.[[#References |[1]]] Species of fungi that reside within living plant tissue without causing symptoms of disease in their host are known as fungal endophytes.[[#References |[4]]] Fungal endophytes colonize a variety of both monocot and eudicot plants which suggests that this symbiosis predated the monocot-dicot split that occurred 140-150 million years ago.[[#References |[2]]] It is also hypothesized that it was phototroph-fungi associations that enabled plants to first colonize land.[[#References |[3]]] This mutualistic association could have helped plants acclimate to new environmental stresses such as desiccation, increased exposure to solar radiation, and more extreme temperatures differences.[[#References |[3]]] | <br>It has been estimated that over 80% of terrestrial plants form a [http://en.wikipedia.org/wiki/Symbiosis symbiotic] association with fungi.[[#References |[1]]] Species of fungi that reside within living plant tissue without causing symptoms of disease in their host are known as fungal [https://microbewiki.kenyon.edu/index.php/Plant_endophyte endophytes].[[#References |[4]]] Fungal endophytes colonize a variety of both [http://en.wikipedia.org/wiki/Monocotyledon monocot] and [http://en.wikipedia.org/wiki/Eudicots eudicot] plants which suggests that this symbiosis predated the monocot-dicot split that occurred 140-150 million years ago.[[#References |[2]]] It is also hypothesized that it was phototroph-fungi associations that enabled plants to first colonize land.[[#References |[3]]] This mutualistic association could have helped plants acclimate to new environmental stresses such as desiccation, increased exposure to solar radiation, and more extreme temperatures differences.[[#References |[3]]] | ||
<br><br>Fungal endophytes remain an important component of today’s terrestrial ecosystems, and many enable their hosts to thrive in harsh environments. Studies show that fungal endophytes can enhance the drought, salt, and soil temperature tolerance of their host plant in addition to increasing resistance to parasitic fungi and herbivores.[[#References |[4]]] With growing concerns about climate change and its effects on agriculture, learning about fungal endophyte conferred drought tolerance has become increasingly important. By influencing plant morphology, development, and physiological and biochemical responses to stress, fungal endophytes can induce mechanisms of drought avoidance, drought tolerance, and drought recovery in their hosts.[[#References |[8]]] | <br><br>Fungal endophytes remain an important component of today’s terrestrial ecosystems, and many enable their hosts to thrive in harsh environments. Studies show that fungal endophytes can enhance the drought, salt, and soil temperature tolerance of their host plant in addition to increasing resistance to parasitic fungi and herbivores.[[#References |[4]]] With growing concerns about climate change and its effects on agriculture, learning about fungal endophyte conferred drought tolerance has become increasingly important. By influencing plant morphology, development, and physiological and biochemical responses to stress, fungal endophytes can induce mechanisms of drought avoidance, [http://en.wikipedia.org/wiki/Drought_tolerance drought tolerance], and drought recovery in their hosts.[[#References |[8]]] | ||
==Classes of Fungal Endophytes== | ==Classes of Fungal Endophytes== | ||
===Clavicipitaceous Endophytes=== | ===Clavicipitaceous Endophytes=== | ||
====Class 1==== | ====Class 1==== | ||
Clavicipitaceous endophytes are associated with grasses.[[#References |[4]]] They typically are found within the plant shoots and form systemic intercellular infections.[[#References |[4]]] While they are often passed down in the seed through vertical transmission, they may also undergo horizontal transmission.[[#References |[4]]] Many class 1 endophytes produce alkaloids to protect their host plant from herbivory by insects and mammals, and studies have shown some class 1 endophytes to confer drought and metal tolerance.[[#References |[5]]][[#References |[8]]] <i>[https://microbewiki.kenyon.edu/index.php/Neotyphodium_coenophialum Neotyphodium coenophialum] </i>, for example, stimulates plants to develop more extensive root systems and longer and thinner root hairs. [[#References |[8]]] | Clavicipitaceous endophytes are [https://microbewiki.kenyon.edu/index.php/Grasses_and_endophytic_fungi associated with grasses].[[#References |[4]]] They typically are found within the plant shoots and form systemic intercellular infections.[[#References |[4]]] While they are often passed down in the seed through [http://en.wikipedia.org/wiki/Vertically_transmitted_infection vertical transmission], they may also undergo [http://en.wikipedia.org/wiki/Horizontal_disease_transmission horizontal transmission].[[#References |[4]]] Many class 1 endophytes produce [http://en.wikipedia.org/wiki/Alkaloid alkaloids] to protect their host plant from herbivory by insects and mammals, and studies have shown some class 1 endophytes to confer drought and metal tolerance.[[#References |[5]]][[#References |[8]]] <i>[https://microbewiki.kenyon.edu/index.php/Neotyphodium_coenophialum Neotyphodium coenophialum] </i>, for example, stimulates plants to develop more extensive root systems and longer and thinner root hairs. [[#References |[8]]] | ||
===Nonclavicipitaceous Endophytes=== | ===Nonclavicipitaceous Endophytes=== | ||
[[File:Jacobs_et_al_(2011).jpeg|thumb|400px|right| [http://www.plantphysiol.org/content/156/2/726/F10.expansion.html Figure 1. Four stages of root colonization by class 2 endophyte <i>P. indica </i>. 1. Extracellular colonization of the root surface 2. The hyphae penetrate the epidermal (RC) and cortical (C) cells. Cell organelles (e.g. nucleus; blue) remain intact and plasma membrane invaginate (dark gray lines inside cells). 3. The plasma membrane still surrounds intracellular hyphae while biotrophically colonized cells die (light gray filling of cells) and organelles are disrupted. 4. Fungal reproduction in RC and C cells. Endodermis cells (E) are not colonized. | [[File:Jacobs_et_al_(2011).jpeg|thumb|400px|right| [http://www.plantphysiol.org/content/156/2/726/F10.expansion.html Figure 1.] Four stages of root colonization by class 2 endophyte <i>P. indica </i>. 1. Extracellular colonization of the root surface 2. The [http://en.wikipedia.org/wiki/Hypha hyphae] penetrate the epidermal (RC) and [http://en.wikipedia.org/wiki/Cortex_(botany) cortical] (C) cells. Cell organelles (e.g. nucleus; blue) remain intact and plasma membrane invaginate (dark gray lines inside cells). 3. The plasma membrane still surrounds intracellular hyphae while biotrophically colonized cells die (light gray filling of cells) and organelles are disrupted. 4. Fungal reproduction in RC and C cells. Endodermis cells (E) are not colonized.[[#References |[13]]] ]] | ||
| Line 29: | Line 29: | ||
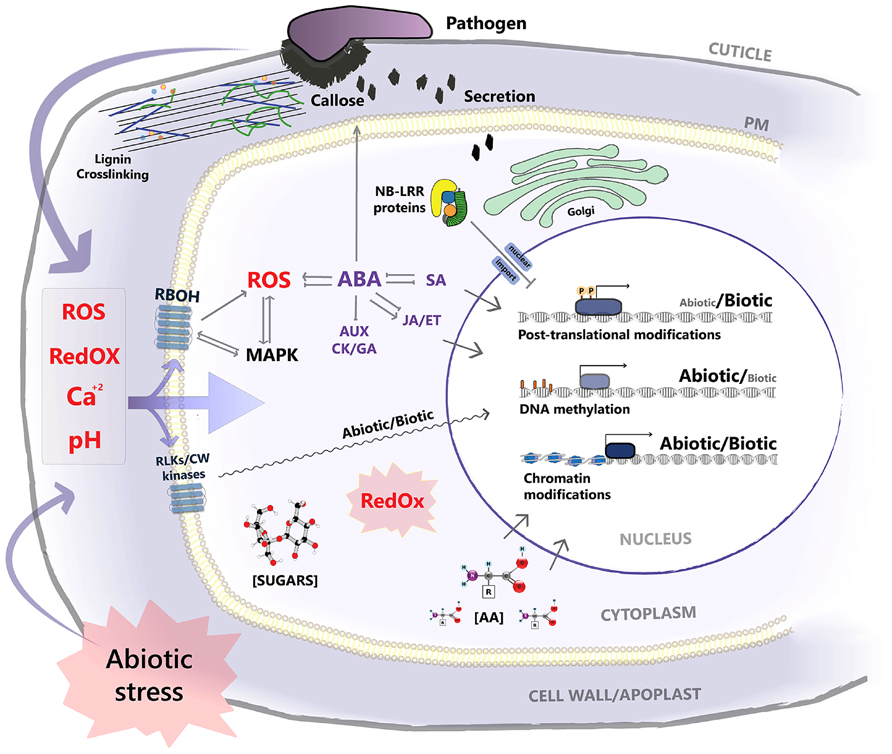
[[File:Kissoudis_et_al_(2014).jpg|thumb|400px|right| [http://www.frontiersin.org/files/Articles/90871/fpls-05-00207-r2/image_m/fpls-05-00207-g001.jpg Figure 2. <b>Signaling pathways of abiotic and biotic stress at the cellular level.</b> Both types of stress factors affect the homeostasis of chemical signals such as Ca2+, ROS, and pH levels. Changes in these levels due to abiotic stress can affect the integrity of physical barriers against pathogens. Both abiotic and biotic stresses also share signaling nodes such as RBOHs (which produce ROS), MAPKs (signal cascades that regulate stress and hormonal responses), RLKs and other cell wall (CW) kinases which are used to regulate gene expression. The stress hormone Abscisic acid (ABA) is central to abiotic stress response. It inhibits hormones responsible for growth and development such as auxin (AUX), cytokinin (CK) and gibberellin (GA) and triggers increased generation of ROS as well as plant defense signaling (JA). Redox state and metabolite concentrations of sugars and amino acids (AA) regulate post-translational modifications. Changes in chromatin and DNA methylation status alter the expression patterns of genes through transcription factor activation and its binding to stress responsive gene promoters. | [[File:Kissoudis_et_al_(2014).jpg|thumb|400px|right| [http://www.frontiersin.org/files/Articles/90871/fpls-05-00207-r2/image_m/fpls-05-00207-g001.jpg Figure 2.] <b>Signaling pathways of abiotic and biotic stress at the cellular level.</b> Both types of stress factors affect the homeostasis of chemical signals such as Ca2+, ROS, and pH levels. Changes in these levels due to abiotic stress can affect the integrity of physical barriers against pathogens. Both abiotic and biotic stresses also share signaling nodes such as RBOHs (which produce ROS), [http://en.wikipedia.org/wiki/Mitogen-activated_protein_kinase MAPKs] (signal cascades that regulate stress and hormonal responses), receptor like kinases (RLKs) and other cell wall (CW) kinases which are used to regulate gene expression. The stress hormone [http://en.wikipedia.org/wiki/Abscisic_acid Abscisic acid] (ABA) is central to abiotic stress response. It inhibits hormones responsible for growth and development such as auxin (AUX), cytokinin (CK) and gibberellin (GA) and triggers increased generation of ROS as well as plant defense signaling ([http://en.wikipedia.org/wiki/Jasmonate JA]). Redox state and metabolite concentrations of sugars and amino acids (AA) regulate post-translational modifications. Changes in chromatin and DNA methylation status alter the expression patterns of genes through transcription factor activation and its binding to stress responsive gene promoters. [[#References |[13]]] ]] | ||
===Osmotic | ===Osmotic Adjustment=== | ||
Osmotic Adjustment is a biochemical mechanism that helps plants to acclimatize to dry and saline conditions. [[#References |[15]] It is an essential process that enables plants to maintain water uptake | Osmotic Adjustment is a biochemical mechanism that helps plants to acclimatize to dry and [http://en.wikipedia.org/wiki/Salinity saline] conditions. [[#References |[15]] It is an essential process that enables plants to maintain water uptake and metabolic processes.[[#References |[15]] Increasing the levels of solutes such as water soluable sugars, prolin, other amino acids, and [http://en.wikipedia.org/wiki/Loline_alkaloid loline alkaloids] leads to a more negative [http://en.wikipedia.org/wiki/Osmotic_pressure osmotic potential] to prevent damage of cell walls.[[#References |[15]] It improves the degree of cell hydration and maintains [http://en.wikipedia.org/wiki/Turgor_pressure turgor] in leaf tissue and in other metabolically active cells.[[#References |[15]] | ||
Redman et al (2015) examined the osmotic concentrations in non symbiotic and heat-stress tolerant symbiotic plants. The pattern was different between the two groups, leading them to conclude that symbiotic plants do not only rely on increasing their osmolyte concentrations.[[#References |[6]]] Endophyte known to promote drought tolerance have high levels of loline alkaloids. [[#References |[7]]] Future experiments could test if these are present in sufficient concentration to prevent the denaturation of macromolecules or reduce the number of reactive oxygen species. [[#References |[7]] | Redman et al (2015) examined the osmotic concentrations in non symbiotic and heat-stress tolerant symbiotic plants. The pattern was different between the two groups, leading them to conclude that symbiotic plants do not only rely on increasing their osmolyte concentrations.[[#References |[6]]] Endophyte known to promote drought tolerance have high levels of loline alkaloids. [[#References |[7]]] Future experiments could test if these are present in sufficient concentration to prevent the denaturation of macromolecules or reduce the number of reactive oxygen species. [[#References |[7]] | ||
===Reactive Oxygen Species=== | ===Reactive Oxygen Species=== | ||
Abiotic stresses such as drought result in the overproduction of [http://en.wikipedia.org/wiki/Reactive_oxygen_species reactive oxygen species] (ROS) [[#References |[10]]] These highly reactive metabolic products act as signaling molecules; however, when their levels are too high, they cause [http://en.wikipedia.org/wiki/Oxidative_stress oxidative stress] and damage proteins, lipids, and DNA. [[#References |[10]]] [[#References |[11]]] ROS control many plant processes such as growth, abiotic stress response, cell cycle, and programmed cell death because they influence the expression of genes. [[#References |[10]]] | [http://en.wikipedia.org/wiki/Abiotic_stress Abiotic stresses] such as drought result in the overproduction of [http://en.wikipedia.org/wiki/Reactive_oxygen_species reactive oxygen species] (ROS) [[#References |[10]]] These highly reactive metabolic products act as signaling molecules; however, when their levels are too high, they cause [http://en.wikipedia.org/wiki/Oxidative_stress oxidative stress] and damage proteins, lipids, and DNA. [[#References |[10]]] [[#References |[11]]] ROS control many plant processes such as growth, abiotic stress response, cell cycle, and programmed cell death because they influence the expression of genes. [[#References |[10]]] | ||
<!-- | <!-- | ||
| Line 76: | Line 76: | ||
[[File:tileshop.fcgi3.jpg|thumb|400px|right| [http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3130040/figure/pone-0014823-g003/ Figure 3. Effects of salt, cold and drought stress and the water usage in S and NS rice plants under laboratory conditions. C) Five week old rice plants (N = 140–210) either NS or S with TempSym1 (TS1) were not watered for the indicated number of days. Control plants (C) were kept hydrated and healthy for both treatments. D) Fluid usage of 5 week old NS or S (SaltSym, TempSym1, and TempSym2) rice plants (N = 60) as ml of fluid used over a ten day period. All three symbionts used less fluid compared to NS plants (Duncan's multiple-range test SD≤7.51; P<0.05). ][[#References |[6]]] ]] | [[File:tileshop.fcgi3.jpg|thumb|400px|right| [http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3130040/figure/pone-0014823-g003/ Figure 3. Effects of salt, cold and drought stress and the water usage in S and NS rice plants under laboratory conditions. C) Five week old rice plants (N = 140–210) either NS or S with TempSym1 (TS1) were not watered for the indicated number of days. Control plants (C) were kept hydrated and healthy for both treatments. D) Fluid usage of 5 week old NS or S (SaltSym, TempSym1, and TempSym2) rice plants (N = 60) as ml of fluid used over a ten day period. All three symbionts used less fluid compared to NS plants (Duncan's multiple-range test SD≤7.51; P<0.05). ][[#References |[6]]] ]] | ||
Redman et al (2015) isolated three Class 2 fungal endophytes from rice relatives to test how well they would confer abiotic stress tolerance to rice plants. <i>Fusarium culmorum</i> (SaltSym) was isolated from the coastal plant <i>Leymus mollis</i> which is exposed to high salt stress, while <i>Curvularia protuberant</i> (TempSym1 and 2) was isolated from <i>Dichanthelium lanuginosum</i> which grows in geothermal soils that can reach temperatures of up to 60 degrees Celcius.[[#References |[6]]] Rice plants with and without the fungal endophytes were grown in greenhouse conditions and subjected to treatments of salt, cold, and drought stress. | Redman et al (2015) isolated three Class 2 fungal endophytes from rice relatives to test how well they would confer abiotic stress tolerance to rice plants. <i>Fusarium culmorum</i> (SaltSym) was isolated from the coastal plant [http://en.wikipedia.org/wiki/Leymus_mollis <i>Leymus mollis</i>] which is exposed to high salt stress, while <i>Curvularia protuberant</i> (TempSym1 and 2) was isolated from [http://en.wikipedia.org/wiki/Dichanthelium_lanuginosum <i>Dichanthelium lanuginosum</i>] which grows in geothermal soils that can reach temperatures of up to 60 degrees Celcius.[[#References |[6]]] Rice plants with and without the fungal endophytes were grown in greenhouse conditions and subjected to treatments of salt, cold, and drought stress. | ||
<br>The study found that rice plants inoculated with the endophytes (S) showed no cost when grown in non-stressful conditions, but the percentage of colonized plants decreased from 100% to 65%.[[#References |[6]]] Plants grown under continual stress, however, maintained 100% colonization.[[#References |[6]]] When infected plants were grown in stressful environments, their water consumption decreased by 20–30% while their growth rate, reproductive yield, and biomass increased (Figure 3). [[#References |[6]]] Non-infected plants (NS), on the other hand, lost shoot and root biomass when exposed to stress. All three endophytes treatments took 2-3 times longer to wilt than the non-colonized plants; however, the mechanism of the conferred drought tolerance remains unknown.[[#References |[6]]] An interesting observation was that the endophytes changed the development of the plants to increase root biomass before shoot growth.[[#References |[6]]] | <br>The study found that rice plants inoculated with the endophytes (S) showed no cost when grown in non-stressful conditions, but the percentage of colonized plants decreased from 100% to 65%.[[#References |[6]]] Plants grown under continual stress, however, maintained 100% colonization.[[#References |[6]]] When infected plants were grown in stressful environments, their water consumption decreased by 20–30% while their growth rate, reproductive yield, and biomass increased (Figure 3). [[#References |[6]]] Non-infected plants (NS), on the other hand, lost shoot and root biomass when exposed to stress. All three endophytes treatments took 2-3 times longer to wilt than the non-colonized plants; however, the mechanism of the conferred drought tolerance remains unknown.[[#References |[6]]] An interesting observation was that the endophytes changed the development of the plants to increase root biomass before shoot growth.[[#References |[6]]] | ||
<br> | <br> | ||
All plant species used in this experiment were members of the family Poacea but belonged to different subfamilies. [[#References |[6]]] The isolated fungal endophytes successfully conferred drought tolerance to the rice plants which supports the idea that the symbiotic communication needed to communicate between the fungi and the plant was conserved within the family. [[#References |[6]]] While many fungal endophytes show habitat-adapted symbiosis, the fact that there is still lower biodiversity in high stress environments indicates that plants require more than just a fungal endophyte in order to be successful. [[#References |[6]]] | All plant species used in this experiment were members of the family [http://en.wikipedia.org/wiki/Poaceae Poacea] but belonged to different subfamilies. [[#References |[6]]] The isolated fungal endophytes successfully conferred drought tolerance to the rice plants which supports the idea that the symbiotic communication needed to communicate between the fungi and the plant was conserved within the family. [[#References |[6]]] While many fungal endophytes show habitat-adapted symbiosis, the fact that there is still lower biodiversity in high stress environments indicates that plants require more than just a fungal endophyte in order to be successful. [[#References |[6]]] | ||
| Line 89: | Line 89: | ||
[http://informahealthcare.com/doi/abs/10.3109/07388551.2013.800018 Khan, A. L., Hussain, J., Al-Harrasi, A., Al-Rawahi, A., & Lee, I. (2015). Endophytic fungi: Resource for gibberellins and crop abiotic stress resistance. Critical Reviews in Biotechnology, 35(1), 62-74. doi:10.3109/07388551.2013.800018] | [http://informahealthcare.com/doi/abs/10.3109/07388551.2013.800018 Khan, A. L., Hussain, J., Al-Harrasi, A., Al-Rawahi, A., & Lee, I. (2015). Endophytic fungi: Resource for gibberellins and crop abiotic stress resistance. Critical Reviews in Biotechnology, 35(1), 62-74. doi:10.3109/07388551.2013.800018] | ||
This review examines gibberellins producing endophytic | This review examines [http://en.wikipedia.org/wiki/Gibberellin gibberellins] producing endophytic | ||
fungi in relation to crops. As fungal endophytes are a "frequently overlooked component of crop | fungi in relation to crops. As fungal endophytes are a "frequently overlooked component of crop | ||
ecology," the authors discuss mechanisms in plant-endophyte stress interactions and suggest areas for future research. | ecology," the authors discuss mechanisms in plant-endophyte stress interactions and suggest areas for future research. | ||
Revision as of 21:13, 14 April 2015
It has been estimated that over 80% of terrestrial plants form a symbiotic association with fungi.[1] Species of fungi that reside within living plant tissue without causing symptoms of disease in their host are known as fungal endophytes.[4] Fungal endophytes colonize a variety of both monocot and eudicot plants which suggests that this symbiosis predated the monocot-dicot split that occurred 140-150 million years ago.[2] It is also hypothesized that it was phototroph-fungi associations that enabled plants to first colonize land.[3] This mutualistic association could have helped plants acclimate to new environmental stresses such as desiccation, increased exposure to solar radiation, and more extreme temperatures differences.[3]
Fungal endophytes remain an important component of today’s terrestrial ecosystems, and many enable their hosts to thrive in harsh environments. Studies show that fungal endophytes can enhance the drought, salt, and soil temperature tolerance of their host plant in addition to increasing resistance to parasitic fungi and herbivores.[4] With growing concerns about climate change and its effects on agriculture, learning about fungal endophyte conferred drought tolerance has become increasingly important. By influencing plant morphology, development, and physiological and biochemical responses to stress, fungal endophytes can induce mechanisms of drought avoidance, drought tolerance, and drought recovery in their hosts.[8]
Classes of Fungal Endophytes
Clavicipitaceous Endophytes
Class 1
Clavicipitaceous endophytes are associated with grasses.[4] They typically are found within the plant shoots and form systemic intercellular infections.[4] While they are often passed down in the seed through vertical transmission, they may also undergo horizontal transmission.[4] Many class 1 endophytes produce alkaloids to protect their host plant from herbivory by insects and mammals, and studies have shown some class 1 endophytes to confer drought and metal tolerance.[5][8] Neotyphodium coenophialum , for example, stimulates plants to develop more extensive root systems and longer and thinner root hairs. [8]
Nonclavicipitaceous Endophytes
.jpeg)
Nonclavicipitaceous endophytes are highly diverse and have been isolated from every major lineage of land plant, which includes nonvascular plants, ferns, conifers, and angiosperms.
Class 2
Class 2 endophytes are usually found in the roots, stem, or leaves of their hosts.[4] Like class 1 endophytes, they can also be transmitted either vertically through the seed coat or horizontally. [4] They are unique in that they can confer habitat-specific stress tolerance to their hosts.[4] They often increase root and/or shoot biomass in their host, and they infect a higher percentage of plants in high-stress environments.[4]
Class 3 and 4
Class 3 colonizes the shoot of plants while Class 4 colonizes plant roots.[4] Both are horizontally transmitted.[4] Class 3 endophytes are highly localized when they colonize a plant, and a diverse number of species can colonize an individual plant.[4] Few studies have been performed on Classes 3 and 4 endophytes, and little is known about their ecological role or their ability to confer tolerance.
Reactions to Drought Stress
.jpg)
Osmotic Adjustment
Osmotic Adjustment is a biochemical mechanism that helps plants to acclimatize to dry and saline conditions. [15 It is an essential process that enables plants to maintain water uptake and metabolic processes.[15 Increasing the levels of solutes such as water soluable sugars, prolin, other amino acids, and loline alkaloids leads to a more negative osmotic potential to prevent damage of cell walls.[15 It improves the degree of cell hydration and maintains turgor in leaf tissue and in other metabolically active cells.[15
Redman et al (2015) examined the osmotic concentrations in non symbiotic and heat-stress tolerant symbiotic plants. The pattern was different between the two groups, leading them to conclude that symbiotic plants do not only rely on increasing their osmolyte concentrations.[6] Endophyte known to promote drought tolerance have high levels of loline alkaloids. [7] Future experiments could test if these are present in sufficient concentration to prevent the denaturation of macromolecules or reduce the number of reactive oxygen species. [7
Reactive Oxygen Species
Abiotic stresses such as drought result in the overproduction of reactive oxygen species (ROS) [10] These highly reactive metabolic products act as signaling molecules; however, when their levels are too high, they cause oxidative stress and damage proteins, lipids, and DNA. [10] [11] ROS control many plant processes such as growth, abiotic stress response, cell cycle, and programmed cell death because they influence the expression of genes. [10]
Testing Endophyte Conferred Tolerances

Redman et al (2015) isolated three Class 2 fungal endophytes from rice relatives to test how well they would confer abiotic stress tolerance to rice plants. Fusarium culmorum (SaltSym) was isolated from the coastal plant Leymus mollis which is exposed to high salt stress, while Curvularia protuberant (TempSym1 and 2) was isolated from Dichanthelium lanuginosum which grows in geothermal soils that can reach temperatures of up to 60 degrees Celcius.[6] Rice plants with and without the fungal endophytes were grown in greenhouse conditions and subjected to treatments of salt, cold, and drought stress.
The study found that rice plants inoculated with the endophytes (S) showed no cost when grown in non-stressful conditions, but the percentage of colonized plants decreased from 100% to 65%.[6] Plants grown under continual stress, however, maintained 100% colonization.[6] When infected plants were grown in stressful environments, their water consumption decreased by 20–30% while their growth rate, reproductive yield, and biomass increased (Figure 3). [6] Non-infected plants (NS), on the other hand, lost shoot and root biomass when exposed to stress. All three endophytes treatments took 2-3 times longer to wilt than the non-colonized plants; however, the mechanism of the conferred drought tolerance remains unknown.[6] An interesting observation was that the endophytes changed the development of the plants to increase root biomass before shoot growth.[6]
All plant species used in this experiment were members of the family Poacea but belonged to different subfamilies. [6] The isolated fungal endophytes successfully conferred drought tolerance to the rice plants which supports the idea that the symbiotic communication needed to communicate between the fungi and the plant was conserved within the family. [6] While many fungal endophytes show habitat-adapted symbiosis, the fact that there is still lower biodiversity in high stress environments indicates that plants require more than just a fungal endophyte in order to be successful. [6]
Further Reading
Fungal endophytes have many potential applications in agriculture and conservation, yet there is still much that is not known about plant-fungi symbiosis and the mechanisms behind it. A more complete understanding of fungi’s role in plant community dynamics and ecosystem stability will inform conservation efforts around the world. Their ability to confer resistance to pathogens and deter insect herbivory also gives them promise as alternative solutions to chemical pesticides and fungicides. Moreover, introducing fungal endophytes to crop may increase their drought tolerance by reducing their water consumption and increasing nutrient uptake. This may be used to increase crop yield in arid climates and mitigate the negative effects of climate change.
This review examines gibberellins producing endophytic fungi in relation to crops. As fungal endophytes are a "frequently overlooked component of crop ecology," the authors discuss mechanisms in plant-endophyte stress interactions and suggest areas for future research.
References
|[1] Smith, S., Read, D., 1997: Mycorrhizal symbiosis, 2nd edn., Academy Press, San Diego.
|[2] Shu-Miaw C., Chien-Chang C., Hsin-Liang C., Wen-Hsiung L. (2004). Dating the Monocot–Dicot Divergence and the Origin of Core Eudicots Using Whole Chloroplast Genomes. Journal of Molecular Evolution. Volume 58, p. 424-441
|[3] Selosse, M., & Le Tacon, F. (1998). The land flora: A phototroph-fungus partnership? Trends in Ecology & Evolution, 13(1), 15-20. doi:http://dx.doi.org/10.1016/S0169-5347(97)01
|[4] Rodriguez, R. J., White Jr, J. F., Arnold, A. E., & Redman, R. S. (2009). Fungal endophytes: Diversity and functional roles. New Phytologist, 182(2), 314-330. doi:10.1111/j.1469-8137.2009.02773.x
|[5] Koulman, A., Lane, G. A., Christensen, M. J., Fraser, K., & Tapper, B. A. (2007). Peramine and other fungal alkaloids are exuded in the guttation fluid of endophyte-infected grasses. Phytochemistry, 68(3), 355-360. doi:http://dx.doi.org/10.1016/j.phytochem.2006.10.012
|[6] Redman, R. S., Kim, Y. O., Woodward, C. J. D. A., Greer, C., Espino, L., Doty, S. L., & Rodriguez, R. J. (2011). Increased fitness of rice plants to abiotic stress via habitat adapted symbiosis: A strategy for mitigating impacts of climate change. Plos One, 6(7), e14823. doi:10.1371/journal.pone.0014823
|[7] Schardl, C. L., Leuchtmann, A. & Spiering, M. J. (2004) Annu. Rev. Plant Biol. 55, 315-340.
|[8] Malinowski DP, Belesky DP. (2000). Adaptations of endophtye-infected cool-season grasses to environmental stresses: mechanisms of drought and mineral stress tolerance. Crop Science 40: 923–940.
|[9] Cheplick GP. (2006). Costs of fungal endophyte infection in Lolium perenne genotypes from eurasia and north africa under extreme resource limitation. Environmental and Experimental Botany 60: 202–210.
|[10] Gill, S, Tuteja N. (2010). Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiology and Biochemistry 48: 909-930
|[11] Gechev, T. S., Van Breusegem, F., Stone, J. M., Denev, I. and Laloi, C. (2006), Reactive oxygen species as signals that modulate plant stress responses and programmed cell death. Bioessays, 28: 1091–1101.
|[12] Jacobs, S., Zechmann, B., Molitor, A., Trujillo, M., Petutschnig, E., Likpa, V., Kogel, K-H., Schaefer, P. (2011). Broad-spectrum suppression of innate immunity is required for colonization of arabidopsis roots by the fungus piriformospora indica. Plant Physiology, 156(2), 726-740. doi:10.1104/pp.111.176446
|[13] Kissoudis, C., van de Wiel, C., Visser, R. G. F., & van der Linden, G. (2014). Enhancing crop resilience to combined abiotic and biotic stress through the dissection of physiological and molecular crosstalk. Frontiers in Plant Science, 5, 207. doi:10.3389/fpls.2014.00207
|[14] Khan, A. L., Hussain, J., Al-Harrasi, A., Al-Rawahi, A., & Lee, I. (2015). Endophytic fungi: Resource for gibberellins and crop abiotic stress resistance. Critical Reviews in Biotechnology, 35(1), 62-74. doi:10.3109/07388551.2013.800018
|[15] Sanders, G., Arndt, S. Osm. 2012. Osmotic Adjustment Under Drought Conditions. Plant Responses to Drought Stress (199-299)
Edited by Sarah Barnes, a student of Nora Sullivan in BIOL168L (Microbiology) in The Keck Science Department of the Claremont Colleges Spring 2014.

{kind=link}