Introduction to Organisms
Introduction
The soil ecosystem in an important environment that allows both plants and animals to survive, grow and reproduce. The significance of the soil as an ecosystem is at times overlooked despite its major contributions to the environment. The soil environment is a complex and varied microbial habitat.[1] The plants and organisms inhabiting the soil contribute to its thriving diversity. Soil life is diverse in morphology, metabolism, size, and many other characteristics. It is believed that “there is sufficient [microbial] DNA in 1 g of soil to extend 1,598km.” [30] Soil is a heterogeneous environment with various zones of rhizosphere, aggregates, organic matter and animal residues.[31] Given each zone is a small component of the larger soil ecosystem, rhizosphere accounts for only 5 to 7 percent of the soil environments but contains over “70 percent of the bacterial- and fungal-feeding nematodes.”[31]
Since, all three domains of life: archaea, bacteria and eukarya, inhabit the soil environment it is difficult to observe each microbe individually within the soil ecosystem.Only a few representative groups of each domain will be discussed to provide examples of the roles these organisms play in soil. This page attempts to outline the major microbial groups within the diverse soil environment.The interactions between soil microbes amongst the soil microbial diversity. The concept of survival of the fittest is seen in the soil food web, which allows only a selective few with the correct nutrients to thrive in a soil environment. The interactions of soil microbes is not only a part of nature but it also only selects those organisms that are fit for survival, growth and reproduction. The soil food web and symbiotic relationships are used to explain the dynamic interactions between soil organisms.
Soil Food Web and Microbial Interactions
To survive in a soil environment, an organism must be able to adapt to changing environments and organism interactions. The interactions seen between organisms allows only the fittest to survive, thrive, grow and reproduce in the soil. The only type of interaction shown in (Figure 1.1) is predation. Other common interactions in the soil environment are:
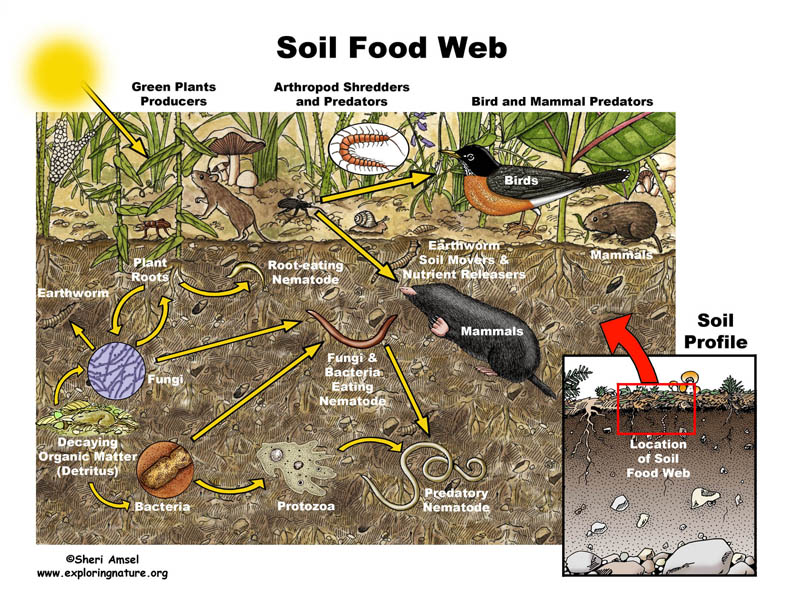
Figure 1.1 Soil Food Web. From[1]

![[1]](http://www.exploringnature.org/graphics/foodwebs/soil_foodweb72.jpg){kind=link}
I. Commensalism: Organism (A) benefits from the presence of Organism (B) while (B) is unaffected by the presence of (A).[32]
II. Mutualism: Organism (A) benefits from Organism (B) and Organism (B) benefits from Organism (A) and becomes a necessary association for each other’s survival. [32] [33]
III. Synergism: Organism (A) benefits from Organism (B) and Organism (B) benefits from Organism (A) but the relationship is not necessary for survival.[33]
IV. Competition: Depending on the organisms this interaction benefits organisms who can readily adapt to the environment and overcome any obstacles needed to survive, grow and reproduce.[33]
V. Amensalism: An interaction in which Organisms (A) and (B) develop a partnership where (B) is unaffected by (A) but organism (A) is negatively affected by (B).[34]
VI. Parasitism: Organism (A) benefits from Organism (B) but Organism (B) does not benefit positively from Organism (A).
VII. Predation: “An interaction between organisms in which one benefits and one is harmed based on the ingestion of the smaller sized organism, the prey, by the larger organism, the predator.”[33]
The soil food web consists of microbes which play an important role in maintaining the complexity of the soil environment. The interactions observed in the food web are not only based on predation, which directly affects the nutrient cycling and degradation of pollutants, but also the formation of aggregates.[35] The production of specific nutrients, such as ammonium created by microbial metabolic functions, helps to enrich the the soil for plant use.[35] Soil organism activity, along with the presence of soil organic matter, create stabilizing aggregates.[35] The hyphae within the soil environment and the microbes help form large, stable aggregates in the soil habitat.[35] The biodiversity in the soil plays a pivotal role in degrading pollutants, and also prevents pathogens from inhabiting the soil environment through competition.[35] The food web in the soil environment is necessary to maintain the soil nutrients, biological activity, and detoxifying pollutants. Thus, it directly maintains cultivation, productiveness and prosperity of the soil.
In (Figure 1.1), all three domains, Archaea, Bacteria, and Eukarya, are represented in the soil food web. If one takes a closer look at the figure, the soil food web is not only dependent on small and large microbes but also dead organic matter and plant roots. Dead and living matter in the soil environment contribute to maintaining the food web. Plant residues also are apart of the soil food web. They contribute to the soil’s carbon source.[35] The plant residues either, release carbon dioxide to the environment, or contribute to the soil organic matter. Soil organic matter contributes to soil’s richness and helps to form aggregates.[35] The main focus in the image above is the diversity of organisms, both in domains and in size. The larger organisms are usually the predators. The smaller organisms eventually fall prey, due to their limited mobility, respiration, and size. The eukarya and small bacteria are apparent figures seen in the figure above, but archaea also contribute to the food cycle as they break down ammonia for plants to use. [35]
The notable interactions of life that take place in the soil environment is evident both below-ground level, as seen in the figure 1.1, but also in the above-ground level.[36] Worms also impact the soil food web, acting as both drivers of the food web both belowground and aboveground.[36]
3 Domains

Carl Woese and the ssRNA

Carl Woese (Figure 1.3) was an American microbiologist famous for pioneering the technique of phylogenetic taxonomy through use of a 16S (for prokaryotic) and 18S (for eukaryotic) ribosomal RNA, also known as small subunit ribosomal RNA [46]. This refined Linnaean classification, taking out the guesswork classification by using a gene that was present in every living organism- the small subunit ribosomal RNA. This method of classification is comprehensive because all organisms possess this form of genetic material. Ssu rRNA is sequenced because the genes are highly conserved. Due to their functional constancy, some have variable regions to differentiate organisms, and are easy to sequence. This technique also elucidated the tree of life, which shows the three domains of microbial lineage diversity: Bacteria, Archaea, and Eukarya. DNA can be compared using a sample database that allows identification of unknown organisms to a species level.
Domain Bacteria
Introduction to Bacteria
Bacteria are single celled prokaryotic organisms that lack membrane bound organelles. They are involved in many important processes within the soil such as decomposition, bioremediation, symbiosis and biological transformation of nutrients. They are present in all terrestrial soil environments.[1] Soil is a heterogenous environment that varies in moisture content, temperature, oxygen requirements, carbon source and many other bacterial growth conditions. Those wide ranging growth factors account for the enormous diversity in soil bacteria populations. One gram of soil can contain between 26,000 to 52,000 different bacterial species.[13] In the entire microbial world, only about 1 percent of bacterial species can be cultured under laboratory conditions which leaves an immense number of bacterial species yet to be described. [14]
Classification based on phenotypic characteristics, such as cellular morphology, metabolism, gram staining, oxygen requirements, carbon source and other traits, was used more readily before modern DNA sequencing. But, phenotypic classification is still used and Bergey’s Manual of Systematic Bacteriology is the most widely used source for determining species based on characteristic species traits.
Traditional species definitions are not as useful in bacteria. OTUs, or operational taxonomic units, based on DNA sequencing similarity, are more applicable to bacteria. Typically, those bacteria with 97 percent similar DNA sequence are classified within the same OTU.[15] The 16S small subunit ribosomal RNA is used to substantiate phylogenetic relationships between different bacterial groups.
Morphology
Bacterial cells measure approximately 1μm wide and 1-5μm long.[12] They are smaller than eukaryotic microbes.Due to their small size, bacteria can more easily adapt to changing soil environments than larger eukaryotes.[1] Most bacteria are singled celled, but actinomycetes grow in branching filaments. There are many different bacterial cell shapes. The most common are bacilli (rods), cocci (spheres), spirilla (spirals) and actinomycete (filaments).

Flagella - There are many different types and different arrangement of flagella. Movement in soil environments is dependent on soil moisture content. Though movement in soil is limited, flagellar function is important to allow bacteria to find more favorable microsites.[1]
Cell membrane - The cell membrane encloses the cytoplasm. It is semi-permeable phospholipid bilayer that regulates transport of nutrients into the cell and waste products out of the cell.
Cell wall - Most bacteria have a rigid cell wall surrounding the cell membrane which protects the cell from osmotic lysis. The cell wall is made of a crosslinked polysaccharide, called peptidoglycan, that is found only in bacteria.[1] The cell wall gives the cell shape and structure, but it is porous enough to permit small water soluble molecules to flow through to the cell membrane surface.[1]
- Gram positive bacteria have a thick cell wall made of several layers of peptidoglycan. The cell wall is the outermost structure, as they do not have an outer membrane. The thick cell wall structure make gram positive bacteria more resilient to osmotic shock. Gram positive bacteria can tolerate 5 to 10 times more turgor pressure than gram Gram negative bacteria.[1] Teichoic acids in the cell wall may also help to bind to cationic nutrients in soil surroundings.[1]
- Gram negative bacteria have a thin cell wall surrounded by an outer membrane made of phospholipids and lipopolysaccharides. Polysaccharides of the outer membrane interact with surrounding cations and are involved in attachment of bacteria to soil particles. [1] Periplasm is a gel like substance that occupies the space between the outer membrane and the cell wall of gram negative bacteria.[1] The outer membrane and periplasmic space is thought to provide protection from environmental toxins.[1] Proteins within the periplasm are thought to begin the process of catalyzing hydrolysis of organic and toxic substances.[1]

Endospores are made by Bacillus and Clostridium in response to environmental stress.[1] Endospores are extremely resistant structures that can withstand extreme temperatures up to 150℃.[12] They can also withstand extreme pressure, desiccation, radiation, pH and chemicals.[12] The endospore protects the bacterial DNA until environmental conditions are more favorable and germination can proceed. Soil is the most common habitat for endospore forming bacteria.[12]
Extracellular polymers - Many bacteria secrete a slime layer or capsule made of polysaccharides or glycoproteins.[1] The outer polysaccharide coating of bacteria helps in attachment to surfaces and the formation of biofilms.[1] Bacterial capsule secretions play a big role in soil aggregation and soil structure.[1] When the carbon to nitrogen ratio within soils is high, microbes secrete the excess carbon as extracellular polysaccharides.[1]
Reproduction
Most bacteria reproduce by binary fission.[12] The bacterial cell expands to twice the original cell size, duplicates the bacterial DNA and pinches off, producing two daughter cells. Each resultant daughter cell is a clone of the parent cell. Prokaryotic Archaea also reproduce by binary fission.
- Spore formation: Actinomycetes are a large group of filamentous bacteria, primarily found in soil.[1] Actinomycetes are morphologically similar to fungi. Actinomycetes form long filaments called hyphae. Unlike most bacteria, the majority of actinomycetes reproduce using asexual spores called conidia.[1] Though they share many characteristic with fungi, actinomycetes are still prokaryotes. They are smaller than fungi and the cell wall contains peptidoglycan.[1] Actinomycetes break down a wide variety of organic compounds and play an important role in organic matter decomposition. [1] Streptomyces, a subgroup within actinomycetes, produce geosmin, which gives soil the characteristic earthy smell.[12]
Metabolism
Along with archaea, bacteria are the most metabolically diverse and employ many different nutritional strategies.[1]
- Heterotrophs - Most bacteria are heterotrophic, obtaining carbon from preformed organic substances. [1]
- Autotrophs - Carbon dioxide serves as the main source of carbon. Some cyanobacteria, nitrifying and sulfur-oxidizing bacteria are autotrophs.[1]
- Organotrophs - Most bacteria obtain electrons from organic substances.[1]
- Lithotrophs - Some cyanobacteria, nitrifying bacteria and sulfur-oxidizing bacteria obtain electrons from inorganic substances.[1]
- Phototrophs - Cyanobacteria use light as the main source of metabolic energy.[1]
- Chemotrophs - All other nonphotosynthetic bacteria do not rely on light for metabolic energy.[1]
Contributions to the Cycle
Bacteria play a significant role in the cycling of a wide range of elements.
Carbon Cycle:
- Carbon Assimilation is the process of fixing carbon dioxide into organic forms. Carbon fixation is performed by autotrophic bacteria, such as cyanobacteria.[1] Chemoautotrophic nitrifying bacteria, such as Nitrobacter, Nitrosospira and Nitrosomonas, also fix carbon.[1] Sulfur oxidizing bacteria, such as Thiobacillus, fix carbon as well.
- Carbon Mineralization results from microbial decomposition of organic carbon compounds into inorganic forms. As the organic carbon substances, like plant matter, are decomposed, inorganic carbon dioxide is produced. Most bacteria are heterotrophs that perform carbon mineralization. When microorganisms die, they are decomposed by successive microbial generations. Peptidoglycan, the main component of bacterial cell walls, is utilized as a polysaccharide substrate by other heterotrophic soil microbes.[1]
- Methanogenesis - Methanogenic bacteria produce methane from the reduction of carbon dioxide. They are all strict anaerobes that use carbon dioxide for the carbon source.[1] They can all use hydrogen as a source of energy and electron donor. [1]
Nitrogen cycle:
- Nitrogen fixation: Azotobacter is a free living nitrogen fixing bacteria.[1] Rhizobium lives in a mutualistic relationship with plants by fixing nitrogen within root the nodules of legumes.[1] Anabaena and Nostoc have specialized heterocyte structures where nitrogen fixation occurs.[1]
- Nitrification: Nitrifying bacteria, such as Nitrobacter, Nitrosospira, and Nitrosomonas, oxidize ammonium to nitrate.[1]
- Denitrification: Alcaligenes, Flavobacterium, and Pseudomonas are common soil denitrifying bacteria.[1]
Domain Archaea
Introduction to Archaea
Vladimir Vernadsky decided, in 1926, that only two domains of life existed on Earth, Bacteria and Eukarya.[37] About 50 years later in 1977, Carl Woese and George Fox theorized that part of the bacterial domain possed multiple dissimilarities from bacteria and recognized it to be another domain of life, now called Archaea.[38] Woese and Fox’s theory regarding the differences of archaea were confirmed in 1990’s through the use of 16S rRNA and 18S rRNA.[38]
Despite the strongly held belief, in the 1990s, that archaea only inhabited extreme environments, recent discoveries have suggested their presence in abundant numbers all throughout the Earth. It is suggested that Archaea play notable roles in maintaining nutrient cycles.[37] Various methods of research, such as cultivation, culture-independent techniques and isotope-based methods, have facilitated the study of domain Archaea in carbon and nitrogen cycling. [37]
The use of genomic and biochemical data has permitted the study of archaeal structure.[37] Use of these research methods has shown that domain archaea and domain bacteria differ in cell wall characteristics yet, share various ways to transfer genetic information by replication and transcription from their common ancestor, Eukarya.[37] As of 2012, 116 genera of Archaea have been observed which consists of 450 species. Most of the archaeal species in these genera remains uncultivated, known only through its gene sequences and molecular surveys.[37]
Morphology
Archaea are characterized by various cell sizes. They are most commonly found to be small organisms ranging from “0.1 to 15 μ diameter and up to 200 μ long.”[38] The shape of many archaea are similar to bacteria, including bacillus, cocci, spirilla and “plate-like forms.” [38] Their cell membranes have ether linked lipids with no periplasmic space, similar to gram-positive bacteria, and contain no intracellular organelles.[39]
Flagella: Most motile archaea inhabit a motor rotary system that allows it to move its flagella through the chemical gradient similar to bacteria.[39] Despite its similarity in function and source of energy required for motility, the flagella of both archaea and bacteria differ in structure.[39] Archaeal flagella gets the energy for its movement by ATP and is composed of dependent filaments that rotate singularly. [40] Archaeal flagella are known to be thinner than bacteria flagellin.[40] Since, the archaeal common ancestor is not bacteria but eukarya, convergent evolution is a plausible explanations for the differences between bacterial and archaeal flagellin. [40]
Membranes: Archaea have a cell membrane that protects the cytoplasm and the nucleic acids from the environment. The archaeal cell membrane has a lipid bilayer with glycol-ether lipids that are more stable than ester linkages and help some archaea that live in extreme environments to withstand such conditions.[39] Most archaeal species are surrounded by a cell wall which protects the cell membrane and maintains the archaeal structure.[41]