Yersinia Pestis and the Plague
Scientific Classification

By Ian Law
Domain: Bacteria
Phylum: Pseudomonadota
Class: Gammaproteobacteria
Order: Enterobacterales
Family: Yersiniaceae
Genus: Yersinia
Species: Y. pestis
Introduction
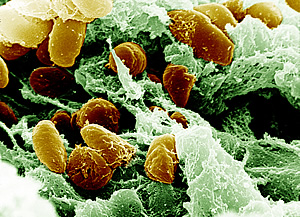
The Yersinia genus comprises various bacteria that cause a spectrum of diseases, spanning from gastroenteritis to the lethal plague caused by Y. pestis. Y. pestis, a gram-negative bacterium, is the pathogen behind the deadly and infectious disease recognized as the plague; shown in Figure 1.[1]. This bacterium holds an intriguing and infamous history, triggering numerous lethal pandemics across human history, notably the Black Death in the 14th century that claimed the lives of millions of people in Europe[1].
Belonging to the family Yersiniaceae, Y. pestis is classified as a facultative anaerobe, capable of thriving in both aerobic and anaerobic conditions. This bacterium uses aerobic respiration and anaerobic fermentation to produce hydrogen gas for energy. Y. pestis has three strains: bubonic, septicemic, and pneumonic. The most frequently occurring form is the bubonic plague, transmitted through an infected flea's bite. The most severe pneumonic form can spread from person to person via respiratory droplets, and it carries unique plasmids that encode essential virulence determinants for the transmission of the plague[2][3].
Y. pestis has been divided into three distinct biovars: Antiqua, Medievalis, and Orientalis. Although they do not differ in virulence or pathology in animals or humans, isolates within a strain exhibit heterogeneity in pulsed-field gel electrophoresis, indicating frequent spontaneous DNA rearrangements, possibly due to insertion sequences in the genome[1].
The evolution of Y. pestis has been rapid, transforming it from an environmental pathogen to a blood-borne pathogen capable of parasitizing insects with limited survival outside a host. Though non-motile, it possesses two distinct flagellar gene clusters[4].
Diagnosis, Treatment, and Prevention
Symptoms
Y. pestis causes three forms of human plague: bubonic, pneumonic, and septicemic, each with varying severity depending on the mode of entry and the bacterium's virulence. To avoid the immune response, Y. pestis utilizes various virulence factors, including F1 antigen, plasminogen activator, and Yersinia outer proteins[5][4]. Bubonic plague, the most common form, is transmitted through the bite of an infected flea or the handling of infected animals. Symptoms typically appear within two to eight days and include fever, headache, chills, nausea, vomiting, abdominal pain, and weakness. The bacterium then spreads through the lymphatic system, causing painful lymphadenitis and the formation of swollen and tender buboes in areas such as the cervical, axillary, and groin regions[4]. The skin overlying the bubo becomes red, warm, and swollen. In some cases, bubonic plague can progress to secondary pneumonic plague (septemic), which is transmitted through respiratory droplets and can be transmitted from person to person[4]. The pneumonic form is characterized by an abrupt onset of fever, headache, weakness, cough, chest pain, and dyspnea and may result in respiratory failure if not treated within 24 hours. Septicemic plague may occur primarily or as a complication of bubonic or pneumonic plague, causing fever, shock, disseminated intravascular coagulation, cyanosis and necrosis, purpura, and gangrene, leading to the disease's nickname, "black death."[4]

Diagnosis
The World Health Organization outlines a set of guidelines for defining a case of plague as suspected, presumptive, or confirmed. These criteria consider clinical symptoms, epidemiological factors, and laboratory results[1][6]. Suspected cases may exhibit symptoms consistent with plague, such as recent travel to an endemic region, contact with infected animals or patients, or flea bites. Presumptive cases meet clinical and epidemiological criteria and specific laboratory criteria, including detection of the Y. pestis bacteria, F1 antigen, serum anti-F1 antibody, or PCR detection of Y. pestis. Confirmed cases meet the same criteria as suspected cases and demonstrate isolation and identification of Y. pestis, along with a four-fold increase in anti-F1 antibody titer in paired serum samples[1][6].
The most prevalent form of the plague is bubonic and is distinguished by symptoms such as fever, chills, headaches, and swollen lymph nodes. Outer extremities would also begin to blacken, pictured in Figure 2. This variant may develop into secondary septicemia, which is accompanied by high mortality rates[6]. On the other hand, primary septicemic plague affects patients with positive blood cultures but no detectable lymphadenopathy[1][6]. In contrast, the primary pneumonic plague is uncommon but fatal and spreads through respiratory droplets. Diagnosis relies on exposure history and symptoms, and laboratory tests are employed to confirm the diagnosis, using blood, bubo aspirates, sputum, cerebrospinal fluid, and skin lesion scrapings as samples. Treatment involves administering antibiotics, while disease prevention entails managing rodents and fleas[6].
To diagnose plague in a laboratory, the standard is to isolate and identify the pathogen from clinical specimens[6]. Various media like agar can be used for culturing, and Y. pestis grows best at temperatures between 26 to 28°C[1]. Detection can also be done through immunological and nucleic acid-based methods. Immunochromatographic assays and portable real-time quantitative PCR thermocyclers have been developed to enable point-of-care testing in remote areas. However, it is important to note that accuracy can only be ensured with proper training[6].
Treatment/Prevention
Effective treatment of Y. pestis is crucial to combat this deadly disease. Timely administration of appropriate antibiotics is vital, as delayed treatment can lead to a fatal outcome[6]. Commonly used antibiotics, such as streptomycin and gentamicin, are effective in treating most cases. Alternatively, a combination of doxycycline, ciprofloxacin, and chloramphenicol may also be used. Physical prevention methods, such as wearing a face mask or covering one's mouth with a jacket, can be employed to prevent transmission[6]. Patients with suspected cases of pneumonic or bubonic plague, including those with secondary pneumonic or septicemic plague, should be isolated. The WHO Expert Committee on Plague recommends antibiotic prophylaxis using tetracycline, streptomycin, and chloramphenicol[6].
Combating and managing the spread of plague requires a variety of tactics, including vaccination, environmental management, and surveillance strategies. However, the current vaccine's effectiveness is limited by frequent adverse reactions and the rapid decline of protective antibodies, requiring frequent booster doses. Additionally, the vaccine does not safeguard against the pneumonic form of the disease, which is a significant worry in bioterrorism readiness[3]. The vaccine is made from Y. pestis organisms grown in synthetic media and inactivated with formaldehyde[7]. To determine the need for booster doses, antibody testing should be performed. After the initial three injections, booster doses should be given as needed every six months due to ongoing exposure[7]. To minimize the risk of infection, suggested measures include removing wild-rodent shelters and food sources near residences, treating pets for fleas, and avoiding direct contact with diseased or deceased rodents. When traveling to areas where plague is prevalent, individuals should avoid contact with wild animals and fleas[7]. Vaccination is advised only for high-risk groups, such as laboratory and field personnel who work with Y. pestis, workers in rural regions with enzootic or epidemic plague, and those whose jobs expose them to wild rodents or rabbits in areas with enzootic plague[7].
As scientists continue to study the plague, they are exploring alternative vaccines, such as ones that relate to the F1 protein[6]. Promising results have been observed in animal models, suggesting that F1 protein could offer significant protection against Y. pestis[6]. However, eradicating plague from endemic regions is challenging due to the pathogen's complex life cycle and multiple potential animal and vector hosts. As such, control strategies focus on early detection and rapid response to outbreaks. These strategies include rodent and flea surveillance, epizoological studies, and treatments involving insecticides and rodenticides[1][6]. Public education campaigns are also crucial to raise awareness about the risks of plague exposure. Despite the challenges, ongoing research and surveillance efforts offer hope for continued progress in combating this deadly disease.
Flea Vector Transmission
The genetic modifications that Y. pestis underwent following its divergence from Y. pseudotuberculosis facilitated biofilm development and its growth in the digestive tract of fleas[1]. The capacity to generate environmental biofilm and the presence of hms genes predates this divergence, and only a few modifications were necessary to adapt the potential to form a biofilm to the flea gut environment[2]. Adding two new plasmids and other discrete changes facilitated the sudden shift to flea-borne transmission[2]. Bacterial biofilm formation is a multifaceted process, and there is still much to learn about it in the Yersinia genus. The application of the biofilm developmental state to enable arthropod-borne transmission represents an innovative function that demonstrates the usefulness of multicellular, adherent growth[2].

Fleas transmit Y. pestis through three different mechanisms[2]. The first is simple mechanical transmission, which occurs when a flea bites a new host after feeding on an infected host[2]. The flea's mouthparts retain residues from the prior infected blood meal, and this residue inoculates the new host. The second mechanism is biological transmission, which depends on bacterial growth in the flea's midgut, followed by regurgitation of the bacterium into the bite site[2]. Y. pestis form large aggregates in the midgut, which can interfere with the valve function of the proventriculus[2]. This valve connects the esophagus and midgut and opens and closes rhythmically during blood feeding. This interference allows for the regurgitation of blood, along with the bacteria from the midgut or proventriculus, back into the bite site. A new transmission mechanism has been recently described in the squirrel flea (see figure 3), which may not require bacterial interference of proventricular function[2]. This novel mechanism may contribute to the vector competence of fleas where the proventricular blockage is infrequent[2].
Antibiotic Resistence
Many studies have found that the Y. pestis bacterium is resistant to a range of antibiotics, including ampicillin, chloramphenicol, kanamycin, streptomycin, spectinomycin, sulfonamides, tetracycline, and minocycline[8][9]. This resistance is thought to be due to the production of various enzymes by the bacterium. Some strains of the bacterium have spontaneously lost their resistance determinants, but a small percentage remains resistant[8]. The resistance genes have been transferred to other strains through conjugation, and it was discovered that plasmid pIP1202 carries these resistance determinants. Despite the challenges posed by antibiotic resistance, ongoing research is dedicated to finding new ways to combat Y. pestis effectively[8][9].

A separate investigation highlighted the emergence of a doxycycline-resistant variation of Y. pestis, referred to as IP2180H, which can transfer resistance to other related bacteria like Y. pseudotuberculosis and E. coli[9]. This suggests that the resistance is associated with a mobile genetic element. Madagascar is the most active region globally for plague, with a high incidence of human cases annually, and it has been observed that multidrug-resistant enterobacteria and plant pathogens may act as reservoirs for antibiotic resistance plasmids, which can transfer to Y. pestis through horizontal gene transfer[9]. Y. pestis strain 16/95 was discovered to exhibit high-level resistance to streptomycin, which was attributed to the presence of a streptomycin phosphotransferase enzyme. This resistance was effectively passed to other bacterial species like Y. pseudotuberculosis, Y. enterocolitica, and E. coli[9]. The strA and strB genes encode an aminoglycoside 3"O-phosphotransferase [APH(3")-Ib] and a 6-O-phosphotransferase [APH(6)-Id], respectively, and they were identified as the resistance determinants. Even though multiple drug-resistant Y. pestis strains have been identified; this bacterium remains a severe threat to public health[9].
The antibiotic resistance of Y. pestis strain 17/95 was investigated in a 2001 study, which found it highly resistant to several antibiotics like those mentioned earlier[9]. The resistance was attributed to the bacterium's production of enzymes or specific proteins. For instance, the resistance to ampicillin was due to a beta-lactamase enzyme, while resistance to chloramphenicol was due to a chloramphenicol acetyltransferase protein[9]. Interestingly, a small percentage of the Y. pestis 17/95 strain lost its resistance determinants spontaneously, leading to the discovery of a new clone named 17/95-I[9]. Moreover, the study revealed that all resistance genes were transferred to avirulent Y. pestis 6/69cN via conjugation. Plasmid DNA analysis from Y. pestis 6/69, 17/95, and 17/95-I showed that strain 17/95 contained fragments corresponding to an additional plasmid, called pIP1202, which is approximately 150,000 bp long. The plasmid was found to be transferable to E. coli K802N and RR1 at frequencies of about 1 ×10-2, and it belongs to the incompatibility group Inc6-C and carries several antibiotic resistance genes[9].
References
[8]
[5]
[9]
[2]
[3]
[4]
[1]
[7]
[6]
- ↑ 1.0 1.1 1.2 1.3 1.4 1.5 1.6 1.7 1.8 1.9 [Perry, Robert D., and Jacqueline D. Fetherston. "Yersinia pestis--etiologic agent of plague." Clinical microbiology reviews 10, no. 1 (1997): 35-66.]
- ↑ 2.00 2.01 2.02 2.03 2.04 2.05 2.06 2.07 2.08 2.09 2.10 [Hinnebusch, B. J., and D. L. Erickson. "Yersinia pestis biofilm in the flea vector and its role in the transmission of plague." Bacterial biofilms (2008): 229-248.]
- ↑ 3.0 3.1 3.2 [Josko, Deborah. "Yersinia pestis: still a plague in the 21st century." Clinical Laboratory Science 17.1 (2004): 25-29.]
- ↑ 4.0 4.1 4.2 4.3 4.4 4.5 [Pechous, Roger D., Vijay Sivaraman, Nikolas M. Stasulli, and William E. Goldman. "Pneumonic plague: the darker side of Yersinia pestis." Trends in microbiology 24, no. 3 (2016): 190-197.]
- ↑ 5.0 5.1 [Easterday, W. Ryan, Kyrre L. Kausrud, Bastiaan Star, Lise Heier, Bradd J. Haley, Vladimir Ageyev, Rita R. Colwell, and Nils Chr Stenseth. "An additional step in the transmission of Yersinia pestis?." The ISME journal 6, no. 2 (2012): 231-236.]
- ↑ 6.00 6.01 6.02 6.03 6.04 6.05 6.06 6.07 6.08 6.09 6.10 6.11 6.12 6.13 [Yang, Ruifu. "Plague: recognition, treatment, and prevention." Journal of clinical microbiology 56, no. 1 (2018): e01519-17.]
- ↑ 7.0 7.1 7.2 7.3 7.4 [“Plague Vaccine.” Centers for Disease Control and Prevention, Centers for Disease Control and Prevention, 11 June 1982, https://www.cdc.gov/mmwr/preview/mmwrhtml/00041848.htm.]
- ↑ 8.0 8.1 8.2 8.3 [Cabanel, Nicolas, Christiane Bouchier, Minoarisoa Rajerison, and Elisabeth Carniel. "Plasmid-mediated doxycycline resistance in a Yersinia pestis strain isolated from a rat." International Journal of Antimicrobial Agents 51, no. 2 (2018): 249-254.]
- ↑ 9.00 9.01 9.02 9.03 9.04 9.05 9.06 9.07 9.08 9.09 9.10 [Guiyoule, Annie, Guy Gerbaud, Carmen Buchrieser, Marc Galimand, Lila Rahalison, Suzanne Chanteau, Patrice Courvalin, and Elisabeth Carniel. "Transferable plasmid-mediated resistance to streptomycin in a clinical isolate of Yersinia pestis." Emerging infectious diseases 7, no. 1 (2001): 43.]

{kind=link}
{kind=link}
![[1]](https://biologydictionary.net/wp-content/uploads/2020/12/antibioticresistance.jpg){kind=link}