Mouth
Introduction

The dark, wet, and warm environment of the mouth, with the occasional meal running through it, makes it an excellent niche for microbes to live. Over the past 40 years, scientists have been arduously working to discover the over 500 different species of bacteria in and around the mouth known today. The mouth is comprised of an oral cavity, which includes the teeth and gums, surrounded by the lips, cheeks, tongue, palate, and throat. Each of these habitats offers differing environmental conditions, and as such, is colonized by a different microbial flora. The oral environment is constantly in flux. From birth to around age 12, when the permanent dentition is complete, the local oral conditions are continuously changing as teeth are shed and new ones erupt [A1]. In addition, environmental factors such as, nutrition, diet, hygiene, smoking, dehydration, and even stress, alter the ecological conditions of mouth. Saliva covers all surfaces and serves various important functions, mechanical and nutritional, (digestive, swallowing, cleansing, lubricative, bactericidal, and excretory) in the oral cavity. It is comprised of various proteins and glycoprotein, of which the main constituents include salivary mucins (approximately 25% of saliva), amylase, IgA, and lysozyme. The typical resting pH (6.5-6.9) is slightly more acidic than stimulating pH (7.0-7.5), but often varies depending on the secretion rate. [A2]. While salivary flora does not necessarily represent the microbial composition of the different components of the mouth, it does impact which microbes can live within the oral cavity, and has recently been the target of research in early disease detection.
Teeth
Description
The second largest biomass of bacteria within the mouth is found on the surface of the tooth enamel, with an estimation of 10^11 organisms per gram wet weight [B1]. This large and diverse population of bacteria is able to reside on the enamel because they form well organized communities known as a biofilm or more commonly supragingival plaque. “Biofilms are analogous to the planet Earth, where the properties of the latter as a whole are determined by the interactions of all of the residents as well as interactions of the populations with inanimate supporting structures” [B2]. The bacteria within the mouth not only have to communicate with all the different species living within the biofilm, but they must obtain a strong adherence to a surface in order to not be washed away by saliva.
The teeth are one of the favored surfaces by bacteria not only because it is a hard surface to adhere to, but the saliva and surface location keeps the oral cavity at warm conditions of 35-36°C, moist, at a neutral to slightly acidic pH due to a bicarbonate ion buffer and bacterial metabolites, and provides nutrients for the bacteria [B3][B4]. The bacteria can also obtain most of its nutritional needs from exogenous nutrients introduced by the host’s diet. These nutrients provide carbohydrates, proteins, and glycoproteins needed for the growth [B3]. The two most common sites where supragingival plaque is found are the approximal and fissure surfaces of the enamel. The approximal surface, between the teeth, is protected from abrasion from salivary flow and oral hygiene, has a rich supply of nutrients from saliva, the host’s diet, microbial products, and gingival crevicular fluid, has low oxygen levels, and has a low redox potential; which allows for predominantly anaerobic bacteria [B3][B4]. The fissures, located on the crowns of the teeth, are more exposed to abrasion, often get food particles packed into crevices, has a high oxygen level, and has a high redox potential, so more diverse range of bacteria reside here [B4].
Who lives there
Microbes present
{kind=link}
Numbered ticks are 11 µM apart.
Gram-stained.
Photograph by Bob Blaylock.
The mouth conditions change throughout the day depending on saliva flow, food, and oxygen levels. This fluctuation of oral conditions causes the population of the biofilm within the mouth to also change because it will affect the bacteria-bacteria interactions. Although studies show that the common microflora on the enamel are constantly changing due to environmental conditions and location, the most common bacteria that are found in the supragingival plaque are: gram positive cocci (Streptococcus mitis, Streptococcus oralis, Streptococcus sanguis, Streptococcus mutans, Streptococcus gordonii, and staphylococcus aureus, staphylococcus epidermidis); gram negative cocci and rods (Veillonella spp., Neisseria sicca, and Fusobacteria); and gram positive rods and filaments (Actinomyces spp., Corynebacterium spp., lactobacilli, and Prevotella spp.) [B5][B6][B2]. These prominent bacteria are responsible for the plaque formation due to their interactions with each other and the tooth surface.
Microbial interactions
- Pellicle formation
In order for the bacteria to form a biofilm first adhere to the tooth’s surface. In the mouth, the tooth forms its own layer of protection known as the acquired pellicle. While protection is its main purpose, it also serves as lubrication for the enamel surface and acts as a semi-permeable membrane [B7]. The pellicle is composed mostly from components of saliva such as proteins, lipids, and carbohydrates, and is never larger than one micrometer thick [B1]. Like supragingival plaque, the acquired pellicle’s composition is believed to differ depending on the region of the mouth because the flow of saliva and the biopolymers that makes up the saliva.
The dental enamel is made of inorganic minerals which mostly consist of calcium and phosphate. The proteins of the saliva bind to the most common calcium phosphates, hydroxyapatite, through an electrostatic interaction. Some of these proteins are the phosphoproteins: statherin, histatin, and proline-rich proteins (PRPs). Although these have been commonly found to form the initial layer of the acquire pellicle, studies have shown that there may be other interactions, such as Van der Waal forces and hydrophobic interactions, that also play an important role in protein-enamel interactions. The second stage of pellicle formation involves protein-protein interactions to build up the layers. Like the biofilm, the pellicle is constantly undergoing modifications depending on the host enzymes and microbial metabolites within the mouth.
The pellicle is able to form a protective layer around the enamel surface because of its many antimicrobial components. Some of these components are: lysozyme, Cystatin, sIgA, and Histatin [B7]. Although the pellicle is initially made to protect the enamel surface, its components also makes a ‘good’ surface for adherence. The early colonizers of the biofilm have receptors that have a high affinity for the PRPs, statherin proteins, α-amylase, and glycoproteins on the pellicle [B8][B9].
- Biofilm formation
The bacteria that live on the tooth surface form multi-specie communities known as a biofilm. A biofilm can be called a community because the bacteria work together to organize themselves in such a way to maintain homeostasis by resisting changes in their environment. Biofilms can resist antimicrobial agents, bacterial enzymes, pH changes to some degree due to the different components of the overall structure of the biofilm and individual contributions each species of bacteria contributes [B3][B6]. Formation of the pellicle on the tooth’s surface gives the other bacteria a face to bind to and eventually leads to the formation of a biofilm. The interactions that allow the bacteria to bind to the pellicle are split up into two stages. When the microbe is still some distance to the pellicle-coated enamel, long-rang physicochemical forces allow for weak, reversible attachment. As the microbe comes closer, molecules called adhesion, on the microbe’s surface, form strong, irreversible interactions with the acquired pellicle. Adhesins are molecular components of the binding structures: pili, fibrils and fimbriae. This process happens through strong-range sterochemical forces. Most oral bacteria have more than one type of adhesin on its surface to allow it to be able to bind to different pellicle sites [B7][B12].
The first bacteria that come in contact with the pellicle surface are known as primary colonizers. These bacteria must be able to withstand high oxygen concentrations and abrasion [B6]. Streptococci make up 80% of the primary colonizers, more specifically S. oralis, S. sanguis, and S. mitis [B7]. As these attach themselves, they in turn provide new surface receptors that adhesin proteins of later colonizers can bind. Some of the most common secondary colonizers are Veilonella spp., Actinomyces naselundii, Prevotella spp., and Neisseria. These earlier colonizers deplete the oxygen within the local environment, and allow for anaerobes to bind to the biofilm [B3][B10]. A notable secondary colonizer is Fusobacterium nucleatum which is important base for tertiary and further layers because it forms a bridge between the early and late colonizers. It has receptors that almost all oral bacteria can bind to. As more bacteria attach themselves and multiply they tend to grow upwards in a columnar fashion, perpendicular to the tooth’s surface [B1][B9]. This dictates the overall population and spatiotemporal formation of the biofilm.
The biofilm is able to withstand environmental pressures because they form a protective outer layer. As the Streptococci and other primary colonizers begin their attachment they utilize sugars mainly sucrose to start polysaccharide production. The polysaccharides are synthesized using enzymes called glycosyltransferases which are located on the cell’s surface. When sucrose is broken down it has two components fructose and glucose, further enzymes and bacterial reactions creates bonds between multiple fructose molecules forming fructan. The same is done with glucose which is called glucan. Glucan and fructan are than brought into the cell and can later be broken down and used as an energy source [B13]. The microbes also use these polysaccharides as a glue-like substance to further secure their attachment to the pellicle-coated enamel and also to form one of the staple components in any mature biofilm, the extracellular matrix [B1][B12]. The matrix provides structural support that allows the microbes to attach in a columnar fashion and also gives the biofilm a strong attachment to the pellicle to prevent it from washing away. One would tend to believe that the extracellular matrix is a solid structure surrounding the cell, but on the contrary it is very open with channels and voids running along it. This is to help the biofilm retain nutrients and water; it also allows them to pass these nutrients in between the channels to other microbes in the community.
- Cell-to-Cell Communication
However, the biofilm is not just a simple community where bacteria can easily bind to nascent surfaces to build upon. The formation of the biofilm is more complex, and involves many interactions between genetically similar and distant species. The biofilm must maintain homeostasis, constant environmental conditions, in order to maintain a healthy mouth. The commensal bacteria compete with their neighbors in order to dictate the conditions of the biofilm [B3]. Through antagonistic and synergistic interactions, the bacteria exhibit the following communication possibilities: coaggregation and coadhesion, metabolic products, gene expression and transformation, quorum sensing, as well as cell-cell signaling are the many ways the bacteria communicate [B6][B9]. Studies are still ongoing to fully understand how the oral microflora is able to from such diverse and complex communities, but it is agreed that communication is essential.
For the early colonizers, who withstand many factors from both the environment and their neighbors, they must have a variety of different receptors and adhesins to perform all their functions. The Streptococcus genus is always one the first and most popular bacteria found at the foundation of the biofilm because they are one of the few bacteria that exhibit intergeneric coaggregation, the ability to bind to both the host and other bacteria. For example, S. gordonii binds to multiple receptors on the acquired pellicle as well as with its neighbors. It binds to salivary agglutinin glycoproteins by the receptor SspA/B, to α-amaylase by AbpA, α-2-3-linked sialic acid termini of O-linked oligosaccharides of glycoconjugates by the protein Has, and also recognizes the Pro-Gln dipeptide of proline-rich proteins [B11]. Other early colonizers like actinomyces, capnoyctophagae, haemophili, prevotellae, propionibacteria, and veillonellae also recognize the components in the acquired pellicle through specific cell surface adherence proteins [B11]. However, studies have shown that individual attachment and growth of monospecies can survive within the mouth.
Bacteria-bacteria interactions increase their affinity for a communal lifestyle. Without coaggregation and coadhesion, the multi-specie biofilms would not be able to form. Coaggregation is the recognition and communication between bacteria in suspension that will clump together to form an aggregate that can bind to the biofilm; while coadhesion is the adhesion of individual bacteria cell in suspension with a cell that is already a member of the biofilm. These interactions are mediated by complementary protein-adhesin and saccharide-receptors [B10]. Coaggregation between streptococci and actinomcyes, initial colonizers, help them bind to the acquired pellicle as well as manipulate spatiotemperoral development of plaque. Cell-cell interactions are formed through lectin-like receptors; which involves a protein adhesion recognizing the streptococcal receptor polyscahharide (RPS). Streptococci can interact between each other; seen when Gn RPS on S. oralis binds with the protein adhesion DL1 on S. gordonii. Coaggregations also occur between different genuses; for example, Gn and G RPS on streptococci is recognized by the protein adhsin type 2 fimbriae on actinomyces [B15]. Without coaggregation, both S. oralis and A. naelundii would not have a high enough affinity to form even a monoculture on the acquired pellicle, but by their combined metabolic products are able to use the salivary components [B2]
Coaggregation and further addition of later colonizers to the biofilm are dictated by the bacteria’s nutritional and environmental needs [B6]. The earlier colonizers are essential to the formation of the biofilm because they change the environmental conditions for the next layer of bacteria to adhere. After S. gordonii has adhered to the acquired pellicle, its interaction with the saliva results in the gene expression of DL1 which encodes for the antigen I/II adhesions, specifically SspA/B. The presence of these proteins promotes its ability to bind to salivary agglutinin as well as the coaggregation of S. gordonii with the other colonizers Actinomyces and P. gingivalis [B8][B9].
Throughout the biofilm, the overall structure is dictated by antagonistic and synergistic interactions between the bacteria. The composition of the biofilm has a basic organizational method, but the actual percentage of each bacterium present differs between individuals most likely because of interactions that occur. A. naeslundii coadheres to S. oralis in an interaction that promotes the growth for both partners. However, since S. gordonii competes for the same nutrients, if A. naeslundii-S. oralis are present, there is a decrease in the S. gordonii population [B10]. The initial colonizers will determine which colonizers will be able to adhere later. For example, if S. mutans is present on the biofilm first, its production of hydrogen peroxide will inhibit the binding and growth of S. mutans since the anaerobe cannot destroy this reactive oxygen intermediate. If S. mutans binds first, its produces bacteriocins, Mutacin I and IV, which will prevent S. sanguinis from binding since it cannot tolerate this toxin [B2][B14]. Some interactions promote synergistic behavior. Some bacteria act as a bridge between bacteria that would not normally coadhere or coaggregate with each other [B9][B2]
One of the most important advantages that bacteria have by living in a biofilm, is that the bacteria are able to withstand more environmental pressures because if the biofilm structure and communication. The bacteria send diffusible signals to their neighbors in order to communicate. One method, quorum sensing, involves the signaling molecule, autoinducer-2 (AI-2) produced from the gene LuxS. This is frequently found in both gram-positive and gram-negative bacteria and is suggested to have a wide range of functions for the community. In biofilms that do not have properly working AI-2 molcules, or those similar, are not able to maintain homeostasis. S. mutans specifically use a competence stimulating peptide (CPS), and the production of this molecule increases its ability to perform transformation by 10-600 times more frequently [B8][B16]. Gene exchange allows the inhabitants the ability share. If a cell lyses, the free DNA can be up taken into a competent cell and incorporated into its own genome, can be used as a nutrient source, or just as a means of communication [B9]. The transfer of conjugative transposons, jumping genes, can allow for horizontal gene transfer, between different geniuses. The bacteria are able to pass genes encoding for antibiotic resistance, such as tetracycline used to treat periodontal disease, as well as “pathogenicity islands” that allow the pathogens to evolve into different strains that can evade past treatments [B9][B16].
Do the microbes change their environment?
- Metabolism
Oral bacterial metabolism is not only important for one species growth and development but important for the entire biofilm as well. For example, Streptococci spp. which is the most prevalent bacteria in the mouth ferments carbohydrates which produce lactic acid. Veillonellae in turn uses the lactic acid produced by Streptococci to promote its own growth. Studies have shown that when Streptococci are absent Veillonellae is not able to grow, so this shows that Veillonellae growth is dependent on Streptococci metabolism [B10].
Metabolism by bacteria in the mouth also produces by products that protect the mouth from incoming pathogens and acid build up. As mentioned already Veillonellae takes up lactic acid which prevents high build up of lactic acid and allowing the mouth to maintain a constant pH. S. oligogermentan has also been shown to use lactic acid and produces a by product of hydrogen peroxide. The elevated levels of hydrogen peroxide inhibit growth of A. actinomycetemcomitans which has been proven to be a big factor in periodontal disease [B2][B6].
- Diseases
Dental plaque is present in both a healthy and diseased mouth, but its only when homeostasis is not maintained that problems arise. Any changes that disrupt the homeostasis of the tooth environment, such as a change in diet (endogenous/exogenous), adhesion, cofactors for growth, pH, redox potential, gases, metabolisms of other biofilm members, abrasion from swallowing, brushing and flossing, the eruption or loss of teeth, drugs, and radiation, can cause the normal microflora species to change. Diseased mouth have a greater number of gram negative cocci, rods, filaments, and anaerobic bacteria [B3][B6]. These changes allow the commensal bacteria to be taken over by the pathogenic bacteria. Now the pathogenic bacteria present in the dental plaque can begin to induce changes in neighboring bacteria gene expression that will allow itself to rapidly increase its own numbers to cause diseases [B3]. An example of this is when metabolic products of the bacteria lead to dental caries, better known as cavities. Today, most host’s diets contain many fermentable carbohydrates which aciduric bacteria, S. mutans and lactobacilli, can easily fermented into organic acids, such as lactic acid. The increase in acids decreases the pH on the teeth. Once the pH has dropped below 5.5 the enamel begins to demineralize. The host’s system tries to counter the demineralization of the tooth surface by having the supersaturated saliva as a buffer system. The saliva is full of calcium and phosphate that is used to re-mineralize minor chips. However, if carbon sources are depleted and the saliva becomes unsaturated, the pH cannot readjust and neutralize leading to tooth decay [B18][B3].
Changes in the supragingival plaque have been shown to influence the subgingival plaque as well. The biofilms in both the supra- and sub-gingival areas have many of the same bacteria with similar interactions. Many of the Streptococci spp., Actionmyces spp.,Prevotella spp., Propionibacterium spp., Veilonella sppl, and F. nucleatum are found in both plaque formations [B11]. According to recent studies, findings have shown that with frequent professional and personal cleaning of the supragingival plaque, there is an efficacy effect on the subgingival plaque and can help reduce a person’s susceptible for diseased periodontal crevices [B17]
Gingiva
Anatomy

The mucus membrane that conceals the alveolar bone, or in other words, the gingiva, is affected by all the environmental and developmental factors which the oral cavity as a whole encounters. The Gingiva is divided into the free gingiva, attached gingiva, and alveolar mucosa by two imaginary lines at the base of the gingival sulcus (also known as gingival crevice) and the mucogingival junction, respectively. The free gingiva is the epithelial tissue from the gingival margin to the imaginary line extending from the cementoenamel junction of the gingival sulcus. Width of the gingival sulcus usually ranges from 0.5 to 2mm, and its depth seldom exceeds 2 to 3mm. Attached gingiva, comprised of the epithelial tissue, supraalveolar connective tissue, and alveolar bone, continues from the imaginary cementoenamel junction of the gingival sulcus to the mucogingival junction. Finally the alveolar mucosal covers the base of the alveolar bone and continues to the floor of the mouth and the vestibule. [C2]
While the gingiva is comprised of numerous components, none is more important than the gingival sulcus, or gingival crevice. Its unique topography, position, and adjacent community of the teeth, serve as a mechanical retention of bacterial plaque, impervious to cleaning by a simple toothbrush [C3]. In addition, the gingival crevice has the lowest oxidation-reduction potential (Eh), as well as a low overall oxygen level, and alkaline pH [C4]. These conditions make the gingival sulcus an excellent environment for anaerobic microbes to inhabit. Furthermore, the gingival crevicular fluid, which flows through the epithelial cells in the gingival sulcus, contributes to the nutrition and immunologic response of the gingival crevice. Stagnation of the gingival crevicular fluid in conjunction with the growth of anaerobic microbes is though to be the determinant in orodental infections [C5]. As such, the relationship between the gingival sulcus and periodontal disease has made the gingival sulcus the primary target of research.
Healthy Gingiva
Healthy gingiva is coral pink in color and harbors a plethora of microbes; most of which are bacteria, but also include archaea. Known bacteria, that have been successfully isolated and characterized, fall under the following categories: gram-positive facultative and anaerobic cocci, gram-positive facultative and anaerobic rods, gram-negative facultative and anerobic rods, gram-negative and facultative cocci, and spiral forms [C6]. Of these categories gram-positive cocci and rods are the most prevalent in the healthy gingival sulcus, specifically streptococcus and actinomyces, respectively [C7].
Diseased Gingiva
To some degree periodontal disease affects approximately 80 percent of the U.S. population, and is one of the most frequent causes of tooth loss. The severity of the disease ranges from simple inflammation of the gums, gingivitis, to a more serious form, periodontitis, which can go as far as bone damage. [C8] The primary cause of this disease is poor dental hygiene, when then results in a cascade of complex molecular interactions involving bacteria in the gingival sulcus and eventual tissue damage. [C9]
On a microbial level the difference between healthy gingiva and periodontal inflicted gingiva is not the specie of microbes present, but rather the ratio of the aerobic and facultative anaerobes to anaerobic microbes. In general, healthy gingiva will possess more aerobic and facultative anaerobes, while anaerobic microbes will predominate in cases of orodental infections [C10]. The exact mechanism of how this transition takes place is unknown, however, there is speculation that environmental factors – pH, oxygen levels, and nutrients – play a substantial role in the composition of the subgingival microflora [C11]. It is known, however, that bacterial colonization, or plaque, of the tooth pellicle is the precursor to periodontal disease. If not addressed, extended exposure of the gingiva to plaque provokes an inflammatory response, also known as gingivitis. Cases prolonged further still advance to the subgingival surface and form pockets of infection. Bacteria in the pockets will induce production of pro-inflammatory cytokines by the host’s immune system, eventually leading to degradation of connective tissues and bone resorption. [C12]
While the abundance of bacterial species found in orodental infections is estimated at around 350, Porphyromonas gingivalis, Actinomyces naeslundii and spirochete Treponema denticola, are three of the more common bacteria.
Porphyromonas Gingivalis
Porphyromonas gingivalis is a gram-negative, anaerobe that can be found along the gingiva, cheek, and tongue. P. gingivalis possesses fimbriae which it uses to adhere to surfaces rich in initial plaque organism such as oral streptococci and antinomyces naeslundii; however it can also bind to other colonizers such as Fusobacterium nucleatum, treponema denticola, and bacteriodes forsythus. P. gingivalis releases a vast array of enzymes, toxic metabolites and cellular constituents, which are often dentrimental to the host in a variety of ways. Proteases are among the proteinases secreted, and are directly involved in the destruction of host tissues and loss of alveolar bone and supporting periodontal tissues, which are characteristic of periodontal disease. However, the role these proteinases play in adhesion, nutrition, and virulence have not been resolved as they are all interdependent. Apart from protein activity, P. gingivalis has a plethora of destructive enzymes and metabolites – phospholipid A (bone resportion), alkaline and acid phosphatases (alveolar bone breakdown), DNase and RNase, sialidase, volatile sulfur compounds, butyrate and propionate (cytotoxins), and indole in ammonia (cytoxins). [C13][C14] P. gingivalis activity is undeniably crucial to the development and advancement of periodontal disease.
Spirochete-Treponema Denticola
Spirochete Treponema denticola is a gram-negative anaerobe associated with subgingival plaque and consequently periodontal disease. T. denticola has the ability to adhere to both host cells and tissues, matrix proteins, collagen, as well as other bacteria, most notably Porphyromonas gingivalis. After adherence to these surfaces its products - peptidases, proteinases, hemolytic and hemagglutinating activities, adhesions, and proteins with pore-forming properties - provoke immune system cells, resulting in cell damage and liberation of harmful factors into the gingiva. [C15][C16] These factors result in gingivitis and eventually periodontitis.
Actinomyces Naeslundii
Actinomyces naeslundii is a facultative anerobic pathogen that contributes to gingivitis, mild periodontitis, and root surface tooth decay [C17]. A. naeslundii has the ability to adhere to the pellicle of a tooth, epithelial cells, collagen, and basement membrane, as well as the capacity to coaggregate with oral bacteria such as Streptococcus, Veillonella, Prevotella, Porphyromonas, Fusobacterium, and Capnocytophaga. The diverse capabilities of A. naeslundii make it a key contributor to dental plaque. To attach to the enamel surface of a tooth it produces fimbriae, which are thinner and shorter than flegellum. A. naeslundii is unique in that it has a multitude of metabolic properties, both aerobic and anaerobic, that give it the flexibility that allows it to survive and contribute to the build up of plaque. Following the consumption of carbohydrates, this bacterium produces organic acids that results in the erosion of the enamel surface due to plaque build up. If the plaque is allowed to remain, an immune response will promote the inflammation of the gingiva, also known as gingivitis. [C18]
Tongue
The microbes that live in the tongue are able to affect the rest of the oral cavities [D2]. Researchers have found that organisms that cause periodontitis and caries also reside in the tongue [D2]. They move from the tongue to the rest of the oral cavities by saliva [D2]. The tongue being the most populated niche in the oral cavity has a great impact to the rest of the oral cavity regions such as the teeth, cheeks and lips.
Physical Conditions

The tongue is a muscle located on the bottom of the oral cavity [D1]. It is broken down into two distinct parts: the tongue dorsum and the lateral tongue [D1]. Each of these surfaces has different physical properties. The tongue dorsum exterior is keratinized, highly papillated and it has an extensive amount of serous glands [D5]. The lateral tongue has a soft non-keratinized exterior [D5]. Due to their physical differences, they are habitats to different species. The tongue is the most populated niche for many microbes due to is high concentration of crypts [D1]. The crypts are many millimeters deep protecting the microbes from mechanical abrasion while providing them with nutrients [D1]. The nutrients found on the crypts come from food, saliva and dead cells [D1]. The crypts lack access to oxygen compare to the mouth, which has a partial of oxygen of about eight mmHg [D1]. Therefore, they can house many different species of anaerobes [D1]. The pH of the tongue is also ideal for the growth of many microbes since is neutral [D1]. The conditions mentioned before are not constant and can change due to smoking, diet, saliva flow and mechanical abrasion (such as tongue cleaning, tooth brushing and chewing).
In diseased tongue: A healthy mouth is moist with saliva, maintained at neutral pH and at temperature 37℃. This is a good environment for many different microbes to grow on. One of this disorders is Glossitis tongue which can lead to discomfort, difficulty with chewing, speaking and swallowing. It may also block airway blockage.Glossitis is also called geographical tongue. They both mean inflammation of the tongue, a condition when the tongue smells and changes color. Papillae are absent and the tongue looks smooth when glossitis happens. The condition varies with the amount of salivation produced. Saliva inhibits the growth and activity of some microbes. Also good oral hygiene reduces the microbial growth and alleviates the infection of the bacterial disease. The infection can also be treated with antibiotics, antifungal medications, or other antimicrobials [D7]. To prevent disease you should avoid irritants (such as hot or spicy foods, alcohol, and tobacco) to minimize the inflammation. Antimicrobial peptides are important contributors to maintaining the balance between health and disease in this complex environment.
Which microbes are present?
Among the many species that live in the tongue are the following: Veillonella atypical, which mostly live in the tongue and saliva. They co-aggregate with many other microbes that are present in the mouth [D1]. F. nucleatum and Porphyromonas gingivalis are located in the tongue [D1]. These microbes are anaerobic and use fermentation as a source of energy; they live in the crypts of the tongue [D1]. Selenomonas spp. and Actinobacillus actinomycetemcomitans are motile and anaerobes [D1]. The microbes that reside in the tongue and are responsible for bad breath are: Porphyromonas gingivalis, Fusobacterium, Prevotella intermedia and Capnocytophaga spp. [D2]. The white coating of the tongue is form by Odontomyces viscous which live in the dorsum of the tongue [D2]. In a study, researchers concluded that 51% percent of patients had bad breath due to tongue coating [D2].
In diseased tongue: There are mainly three types of microbes living on surface of tongue and play an important role in Glossitis tongue. Infection by Herpes simplex virus type (HSV) 1 and 2 are believed as the virus that causes Glossitis Tongue [D7]. HSV type 1 is always linked with this infection and mostly happened in childhood [D8]. HSV-1 often causes lesions inside the mouth and is transmitted by contact with infected saliva [D8].Another type is Streptococcus family. Streptococcus salivarius [D9], Streptococcus mutans, Streptococcus gordonii and Streptococcus sobrinus[D10] are three of them usually found in human tongue. The third type is Candida albicans species which includes C. glabrata and C. tropicalis yeasts [D11]. Yeast population can vary with the amount of salivation produced in mouth since saliva served as a barrier for external and internal environment [D12].
Do the microbes that are present interact with each other and the environment?
““… the community is capable of doing things that individual (species) cannot” David Relman microbiologist at Stanford University in California [D4]. Most of the bacteria that live in the tongue survive by co-aggregation with other bacteria [D1]. Microbes form proteins that allow other microbes to colonize on top of them [D4]. Many of the bacteria that are present in the tongue are not motile therefore they have to work together to acquire nutrients and energy [D1]. They acquire energy by fermentation of peptides and amino acids, which are naturally located in the crypts [D1]. Microbes found in the tongue can also produce sulfur compounds that are responsible for bad breath [D2]. Bad breath known as Halitosis is not harmful to the host but it does have a great impact on the social and psychological aspects of the individual [D6].
In diseased tongue: Communications can be made among genetically unrelated oral bacteria and DNA can be exchanged in different domains through transformation and conjugation. The interaction between Streptococcus gordonii and Candida albicans is best described as in the following: “Streptococcus gordonii DL1 adheres to Candida albicans via a complex series of interactions. Inactivation of genes that encode the large cell wall-associated proteins CshA and CshB partially inhibits S. gordonii DK1 from adhering to the yeast cells. Further, inactivation of the gene encoding the surface proteins SspA and SspB increases the effect twofold and nearly obliterates adherence.” [D9] Herpes simplex virus would transform its DNA into the cell for initiation of infection. The transformation helps the reproduction of more Herpes simplex virus without using its own nutrients and ATP. “HSV is a nuclear replicating, icosahedral, enveloped DNA virus. The HSV envelope contains at least 8 glycoproteins. The matrix or tegument which contacts both the envelope and the capsid contains at least 15-20 proteins.” Streptococci are Gram-positive spherical bacteria [D13]. Gram positive bacteria have a thick cell call enclose the cell membrane and cell cytoplasm. The thick cell wall served as a barrier for the outercellular and intercellular environment. Also, there are a lot of transport protein that helps exchange nutrients and waste. For Candida albicans species, unfavorable condition causes them to form hyphae and thereby penetrate deeper into the mucosal membrane, causing the tongue inflammation [D11].
Throat
Description

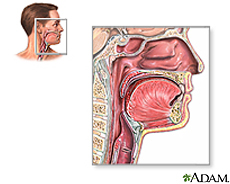{kind=link}
The “end” of the mouth can be considered the pharynx, or more commonly known as the throat. Located behind the mouth and under the nasal cavity, the pharynx is comprised of three parts: nasopharynx (upper), oropharynx (middle), and hypopharynx (lower). [E1] The throat is a muscular passageway which carries food and liquids toward the digestive tract as well as air to the lungs. Because it encounters many different kinds of objects, it’s susceptible to encounters with many organisms. The throat is composed of cells that are able to produce mucus constructing a mucosal membrane with “hairlike projections” (cilia) [E1]. The purpose of the mucus is to facilitate food passage, but is also a way for microbes to latch on to epithelial cells and colonize. The throat is an excellent environment for microbes. Warm and moist, these ideal conditions provide aerobes and anaerobes an environment to flourish. But the throat also offers special atmospheric conditions such as higher exposure to oxygen and carbon dioxide during inhalation and exhalation for microbes which reside there [E2]. Coupled with the plentiful supply of nutrients from eating and breathing (plenty of oxygen), the micro environment in the throat is excellent for microbe growth [E2].
Who lives there?
Which microbes are present?
Both gram-positive and gram-negative bacteria are commonly found in the throat, including a specie of fungi (Candida). The predominant microbes found in the throat are alpha-hemolytic streptococci (viridans streptococci). Viridans streptococci, including species within the S salivarius, S mutans, and S anginosus groups, constitute the most prevalent components of the normal oropharyngeal flora [E6].
The microbes found in the oropharynx area of the tongue are predominantly the viridans group (gram-positive cocci), gram-positive bacilli (corynebacterium), gram-negative cocci (moraxella and Neisseria spp), gram negative bacilli (Haemophilus influenzae-2 and Haemophilus parainfluenzae-3) and gram-negative bacilli (fusobacterium-4 and porphyromonas, prevotella) [E7]. Gram-negative diplococci, including N meningitidis, are also found in 5% to 10% of healthy asymptomatic individuals [E8][E9].
Of particular interest, are some microbes that are found in the oropharynx that are not found in the mouth (saliva and tooth surfaces). These microbes include Streptococcus pyogenes, Streptococcus pneumoniae, Haemophilus influenzae, and Haemophilus parainfluenzae. The first two are gram-positive cocci and the last two are gram-negative bacilli. Of these four microbes, 3 of them are pathogenic. Haemophilus parainfluenzae becomes pathogenic when the conditions warrant it. On the other hand, microbes such as Streptococcus faecalis (gram-positive cocci), Eikenella and enterobacteriaceae (gram-negative bacilli), actinomyces and lactobacillus (gram-positive bacilli), veillonella (gram-negative cocci), and treponema (spirochete) are all found in the saliva and tooth surface, but not in the oropharynx [E10]. The tongue, which is the closest subniche to the throat, consists of microbes that are shared with the throat and microbes that are exclusively on the tongue. For example, S. salivarius is found both on the tongue and throat. However, Veillonella is exclusively on the dorsum of the tongue and the buccal mucosa [E10]. The oral flora develops from the very beginning right after childbirth. Humans are born with a sterile mouth within two days of birth. Streptococcal species, predominantly S mitis and S salivarius, become established and predominate during the first year of life, accounting for 70% of the total flora. [E11][E12].
Do the microbes that are present interact with each other?
The Veillonella species (gram-negative cocci) isolated from subgingival dental plaques demonstrate the ability to coaggregate with streptococcal species, a property vital to the formation of diverse biofilms, whereas those from the tongue and saliva have limited coaggregation properties [E13][E14]. Also, Veillonella are unable to ferment sugars directly and instead rely on fermentation products, such as lactic acid, produced by streptococci. This close relationship with streptococcal species may play a contributory role in the pathogenesis of dental caries [E15].
Within the throat there are hundreds of coexisting and competing microbes. Of the hundreds, Streptococcus pneumoniae and Haemophilus influenzae are the most prevalent [E4]. However when isolated from the same originating specimen, either S. pneumoniae or H. influenzae is more prevalent than the other, or both are significantly less prevalent, which could suggest that there are inhibitory effects of one on the other microbe [E4]. Looking closer at each microbe, it’s understood that both microbes use similar pathways for pathogenesis and colonization within the mucosal layers of the throat, which could explain the decreased prevalence of either microbe when isolated. S. pneumoniae and H. influenzae compete for the same host cell receptor, and thus when antibodies are produced, they tend to wipe out both species [E4]. But the host is not the only factor regulating populations of microbes. Streptococcus pneumoniae has the ability to produce hydrogen peroxide as an inhibitor [E4]. Because the environment of the throat is expected to have high oxygen and increased carbon dioxide, S. pneumoniae’s ability to create hydrogen peroxide is directly related with the amount of oxygen and carbon dioxide present in the environment [E4]. This mechanism has been studied to be an effective way for “limiting and eliminating competitive flora, including H. influenzae and N. meningitides,” which all resides in the same microenvironment (throat).
Do the microbes change their environment?
The microbes have to penetrate the mucosa, which acts as a physical barrier in the throat in order to disrupt the host’s natural defense mechanisms. This layer consists of interconnected epithelial cells resting on a basal membrane. Interspersed with the epithelial cells are mucin-producing goblet cells that line the surface of the nasal cavity and the oropharynx. These cells produce a thick layer of mucin glycoproteins that coat the surrounding epithelia, forming a protective glycocalyx. This layer may trap and block bacteria from adhering to the underlying epithelial cells [E16].
Group A Streptococcus, GAS, is the most common bacteria that cause pharyngitis in the throat. The pathogenesis of GAS infection starts with the attachment to the epithelial cells of the pharynx via hairlike fimbriae that are present on the surface of the cell. Surface virulence factors on the GAS and secreted virulence factors help GAS to overcome the mucosal layer of the epithelial cells lining the throat. On the surface of the GAS, fimbriae are composed of lipotechoic acid and a series of distinct surface proteins, M proteins. These M proteins have important antiphagocytic properties. GAS also have surface molecules called adhesions, for example antigen I/II, that bind to the host receptor proteins such as salivary proteins containing sialic acid moieties on the epithelial cells [E17][E18]. Additionally, host molecules such as fibronectin are associated with these adhesions as well. More specifically, many of the streptococcal species in the throat produce an array of surface molecules collectively known as ‘‘microbial surface components recognizing adhesive matrix molecules’’ (MSCRAMM), which target fibronectin and other molecules for binding to host cells [E19].
The host epithelial cells have mechanisms of their own to defend their mucosal layer. The epithelial cells of the oral cavity and salivary glands have an innate system for recognizing pathogenic microbes by the activation of toll-like receptors (TLRs). TLRs are transmembrane proteins, of which 10 members currently are known to exist in humans. TLRs function as pathogen-recognition receptors that interact with conserved domains on micro-organisms (so-called ‘‘pathogen-associated molecular patterns,’’ PAMPS) [E20][E21]. These PAMPS include protein motifs (TLR4, -5), lipids or lipoproteins (TLR1, -2), and nucleic acid sequences (TLR9) [E22]. Activation of TLRs leads to a cascade of signaling pathways, ultimately leading to the nuclear translocation of nuclear factor kappa B and the transcription and increased production of various proinflammatory or anti-inflammatory cytokines and chemokines that are critical for nonspecific defense in the head and neck region, including the throat and mouth regions [E23][E24].
Conclusion
The mouth is comprised of several subniches that each function according to their different environmental conditions and location. The throat and tongue epithelial cells are constantly shedding, creating a complex environment that bacteria must adapt to in order to form, let alone maintain a community. The throat, tongue, and tooth surfaces constantly deal with saliva being washed over the surface, but the gingival crevices and crypts are protected. However, even with all these differences they all share some similarities. The mucosa covered surfaces are prime targets for bacteria to adhere, so bacteria are found on the throat, tongue, teeth, and gums no matter how healthy someone tries to be. Some of the common genuses of bacteria throughout the mouth are Streptococci, Neisseria, Fusobacterium, Prevotella, and other anaerobic bacteria. These species are able to reside throughout the different locations of the mouth by forming intra- and inter-generic coaggregations. The mouth is subjected to environmental changes, and any disturbance to the conditions of the mouth lead to changes in the microflora. When conditions are disrupted, and the commensal bacteria are outnumbered by pathogenic bacteria, diseases occur. Some of the same bacteria that cause bad breath on the tongue will cause periodontitis disease between the gum and the tooth surface. Although each subniche is its own continent governed by different rules, their actions and inhabitants affect the whole mouth community and is one of the most widely researched niches to understand microflora.
Current Research
1. Correlation between periodontitis and coronary heart disease
Several theories exist to explain the relationship between coronary heart disease and periodontitis. One such theory includes the role of fibrinogen in linking the two. Fibrinogen is a plasma glycoprotein synthesized in the liver. After coagulation, it is converted in Fibrin, the protein involved in blood clots. The study took 95 subjects and separated them into a healthy control group, moderately and severely chronic periodontitis group, coronary heart disease group, and moderately and severely chronic periodontitis coexisting with coronary heart disease group. After measuring routinely diagnosis procedure for both periodontitis and cardiovascular heart disease along with levels of fibrinogen, results showed fibrinogen levels in subjects with MSP and MSP+CHD to be much higher than those of the healthy controlled group. It was concluded that people with periodontitis may be more at risk for developing cardiac heart disease when taking into consideration fibrinogen as the biological basis. [C19]
2. Bacterial markers of periodontal diseases
Among the various bacteria present in the subgingival deposit, three have been the center of study due to their prevalence in periodontal disease. First, the Tannerella forsythensis, an anaerobic Gram-negative known to possess pathogenic potential. Second, the Actinobacillus actinomycetemcomitans with the ability to attach itself to the enamel surface after the consumption of sugar to produce an acid resulting in the erosion of the enamel surface. And the third, the Porphyronmonas gingivalis, a gram negative bacteria with a virulence factor allowing it to invade the gum tissue. They measure the bacteria on 495 subjects aged 6-82 with the use of the 16S ribosomal RNA based polymerase chain reaction (PCR). Their studies showed an increase in the amount of T. forsythensis and P. gingivalis on older subjects when compared to the adolescents. Furthermore, an increase in T. forsythensis was also seen within the tobacco smokers or the subjects. It was finally concluded that the T. forsythensia had a correlation between periodontal disease in tobacco smoking and elder individuals. [C20]
3. Saliva used as Biomarkers for early disease detection
Current research is being done in the mouth to help detect diseases throughout the entire body. As of now, some hard hitting diseases: cancer, cardiovascular diseases, metabolic and neurological deficits are tough to diagnose especially early on. The research is aimed at providing scientists and doctors a simple, inexpensive way to diagnose these diseases through finding disease-associated proteins and genetic markers in saliva. Saliva dectection is already being used to detect HIV, hepatitis A, B, C, and is used to monitor some drugs like marijuana. The NIDCR is currently working on finding the body’s disease-associated salivary biomarkers for those hard to diagnose diseases than developing an accurate, easy to use diagnostic method [B19].
4. Fluorinated food
For a while now some researchers have been looking into fluorinating food sources. This is done with the thought that because fluoride is the best way of preventing big plaque build up on the tooth, since plaque is linked with gingivitis, periodontal disease, and other oral disease. This current research finds that fluorinating food in populations where fluoride toothpaste and daily brushing has almost no additional beneficial effects. The big reason in wanting to fluorinate food is too possibly fight oral diseases in other countries in which brushing is not practiced regularly. This experiment shows that without the brushing, fluorinated foods have a small but significant effect on controlling plaque it also shows that fluorinated foods should not be substituted for brushing with fluorinated toothpaste [B20].
5. Impact of different tongue cleaning methods on the bacterial load of the tongue dorsum
The objective of this research was to observe the impact of tongue cleaners on the amount of microbes present on the tongue. The researchers found out that cleaning only decreases the amount of bacteria on the tongue if is done regularly. If it tongue cleaners are used sporadically then the results are not long lasting. [D3]
6. Paster’s and colleagues research on the microbes responsible for bad breath
Bacteria that live on the tongue are responsible for bad breath. Paster and his colleagues did this research in the year 2003. The research involved six healthy people and five people with bad breath. They found six organisms that are responsible for bad breath. They treated the bad breath with scraping of the tongue and brushing their teeth with zinc that caused a change on the amount of microbes that live in the tongue. [D4]
7. Diversity of Bacterial Populations on the Tongue Dorsa of Patients with Halitosis and Healthy Patients
This study was concentrated in comparing the microbial load of the tongue dorsa of healthy patient and patients with Halitosis. The researches accumulated their data by using culture-independent molecular methods. They use those methods because some of the bacteria that live in the tongue dorsa cannot be cultivated in the lab. Their goal was to be able to classify all the different species that live in the mouth including the not-yet-cultivated species. They perform tongue scrapping of the patients and then analyzed the 16S rRNA genes from their DNA using a PCR. Due to their experiment, they were able to increase the amount of identified species to 600. [D6]
8. Defining the Normal Bacterial Flora of the Oral Cavity
The purpose of this experiment was to classify the bacteria that are found in the tongue through culture-independent molecular techniques. This experiment was also aiming to figure out the different locations where bacterial flora is present within the mouth. They use the amplification of 16s rRNA genes by PCR and purification of PCR products. Their results showed that different sites of the mouth have different types of bacteria depending on the structure of the surface. The main goal is to understand and know all the bacteria present in a healthy mouth so we can have a greater understanding of disease causing bacteria. [D5]
9. Potential role in the prevention of streptococcal infections for Glossitis
A streptococcus is a species commonly survived in human mouth and produce bacteriocin-like inhibitory substances (BLIS). The research aims to study the practical application of BLIS-producing S. salivarius, “relative to the distribution amongst oral streptococci of the genetic loci encoding the lantibiotics streptococcin A-FF22 (SAFF-22) and salivaricin A (SalA).” The paper suggests a few key points.
For lantibiotic SA-FF22, the paper suggests that SA-FF22 homologues is produced by various S. salivarius and that is called salivaricin G32 (SalG32). Also, “Each producer of SA-FF22-like activity displayed across-immunity to the inhibitory products of the other members of the SA-FF22 lantibiotic cluster.” On the other hand, biologically active lantibiotic SalA is is “detected in some strains of S. salivarius.” Also, “anti-S. pyogenes inhibitory activity is produced when SalA-positive S. salivarius are grown in saliva in an in vitro test system.” Finally, S. pyogenes in the oral cavity can be used to develop as an oral probiotics. The resistance to SalA in a sample of alpha-haemolytic cocci is higher than the microbe which is naturally colonized with SalA-producing S. salivarius. This implies that the level of SalA is associated in the oral cavity for a relatively resistant purpose. [D14]
10. Mutational analysis of the group A streptococcal operon encoding streptolysin S and its virulence role in invasive infection
In the article the authors asked which genes encoded the virulence factors that are responsible for the pathogenicity of GAS. In order to determine the genes involved, the authors used inframe allelic exchange mutagenesis and single-gene complementation to answer their question. They determined that nine genes (sag A- sag I) were necessary and sufficient for Streptolysin S (SLS) production. SLS produces the beta-haemolytic phenotype observed for GAS. Furthermore, SLS has structural similarities to other Gram-positive bacterial bacteriocins, a family of molecules which are toxic to other microbes and host cells in the throat. Thus, SLS production may help GAS compete against other microbes for a niche in the epithelial layer. In concluding, SLS helped impair phagocytic activity and both M proteins and streptolysin O (both virulence factors of GAS) collaborate with SLS to enhance epithelial cell cytotoxicity.
11. Reduction in resistance of clinical isolates of Streptococcus pneumoniae
In a recent study, clinical isolates from 2003 and 2005 were compared to samples from 2007. In 2003 and 2005 isolates, particularly in Asia, it was reported that highly pencillin-resistance strains of Streptococcus pneumoniae were beginning to appear. It was thought that the resistance occurred due to gene mutations in the pbp gene sequence, which coded intermediate to full resistance to penicillin antibiotics. Streptococcus pneumoniae have potential to obtain the pbp protein mutation for resistance through intraspecific lateral gene transfers with already resistant strains. In 2007 clinical isolates however, there was a decrease in pencillin resistance in S. pneumoniae, and was linked to reduced sales of oral beta-lactams at pharmacies. However it was later concluded that the decrease in penicillin resistance from reduced sales of oral beta-lactams at pharmacies was because the beta-lactams caused pencillin resistant phenotypes, and not pbp mutations which was previously thought. Pbp mutations are still believed to be related to pencillin resistance, but in the sense as a recovery mechanism for S. pneumoniae to gain macrolide resistance genes.
References
[A1] Skinner F.A. and Carr J.G. The Normal Microbial Flora of Man. London: Academic Press Inc, 1974
[A2] Wilson M. Bacteriology of Humans: An Ecological Perspective. MA: Blackwell Publishing LTD, 2008.
[B1] Gibbons, R. and Houte, J. “Bacterial Adherence in Oral Microbial Ecology.” Annual Reviews. Microbiology. 29 (1975):19-42
[B2] Kuramitsu, H. et. al. “Interspecies Interactions within Oral Microbial Communities.” Microbiology and Molecular Biology Reviews. Dec. 2007. 653-670.
[B3] Olsen, I. “New Principles in ecological regulation – features from the oral cavity.” Microbiology ecology in health and disease. 18 (2006):26-31
[B4] Wilson, M. Bacteriology of Humans: an ecological perspective. Massachusetts: Blackwell Publishing Ltd, 2008
[B5] Rozkiewicz D, et. al. “Bacterial composition in the supragingival plaques of children with and without dental caries.” Advances in Medical Sciences. Vol.51. (2006). Suppl. 1
[B6] Sbordone, L. and Bortolaia, C. Oral microbial biofilms and plaque-related diseases: microbial communities and their role in the shift from oral health to disease. Clin Oral Invest. 2003. 7:181-188
[B7] Hannig, M. Structure and Function of acquired pellicle. Duckworth (ed.) Teeth and their environment. Karger. Switzerland. 2006
[B8] Jenkinson and Lamont. Oral microbial communities in sickness and in health. Trends in Microbiology. Vol 13. No. 12. 12/2005
[B9] Kolenbrander, P. et al. Communication among oral bacteria. Microbiology and molecular biology reviews vol. 66. No.3. sept. 2002. Pg. 486-505
[B10] Kolenbrander, P. Oral Microbial Communities: Biofilms, Interactions, and Genetic Systems. Annual Reviews Microbiology 2000. 54: 413-37
[B11] Kolenbrander, P. and London J. Adhere today, here tomorrow: oral bacterial adherence. Journal of Bacteriology, June 1993, 3247-3252
[B12] Marsh. P. Dental Plaque as a microbial biofilm. Caries Research. 2004; 38:204-211
[B13] Burnett and Schuster. Oral microbiology and infectious disease. The Williams & Wilkins Company, MD. 1978
[B14] Kreth, J. et. al. Competition and Coexistence between Streptococcus mutans and Streptococcus sanguinis in the Dental Biofilm. Journal of Bacteriology. 11/2005; 187(21):7193-7203
[B15] Palmer, R. Coaggregation-Mediated Interactions of Streptococci and Actinomyces Detected in Initial Human Dental Plaque. Journal of Bacteriology. 2003 June. 185(11):3400-3409.
[B16] Marsh, P. Dental Plaque: biological signfcance of biofilm and community life-style. J Clin Periodontol 2005;32:7-15
[B17] Hellstorm, M-K, et. al. The affect of supragingival plaque control on the subgingival microflora in human periodontitis. J Clin Periodontal. 1996. 23: 934-940
[B18] Palmer, R. Jr. et. al. Biocomplexity in the oral cavity – the basics of structure in supragingival bacterial communities. Biofilms. 2004; 1.329-335
[B19] Wong, B. and Segal, A. Salivary diagnostics: enhancing disease detection and making medicine better. European Journal of Dental Education. Vol.12.1.Feb. 2008. Pages 22-29
[B20] Meyer-Luekel, H., Bitter, K., and Kielbassa, A.M. Effect of a Flouridated Food Item on Enamel in situ. Caries Research. 2007.41:350-357
[C1] “Tooth Anatomy.” Gum Disease In-Depth Report. 1997-2008. 26 Aug. 2008. http://health.nytimes.com/health/guides/disease/periodontitis/print.html
[C2] Goldmam Henry M. and Cohen D. Walter. Periodontal therapy, Fourth edition. Saint Louis: The C.V. Mosby Company, 1968.
[C3] Kelstrup Jens and Theilade Else. “Microbes and periodontal disease.” Journal of Clinical Periodontology 1 (1974): 15-35.
[C4] Wilson M. Bacteriology of Humans: An Ecological Perspective. MA: Blackwell Publishing LTD, 2008.
[C5] Ruby John and Goldner Morris. “Nature of Symbiosis in Oral Disease.” Journal of Dental Research 86:1 (2007): 8-11.
[C6] Skinner F.A. and Carr J.G. The Normal Microbial Flora of Man. London: Academic Press Inc, 1974.
[C7] Kesic Ljiljana, et al. “Normal Oral Flora – A New Classifcation – Part I” ACTA Stomatologica Naissi. 2003. University of Nis. 25 Aug. 2008 http://www.medfak.ni.ac.yu/asn/BROJ-41/6-full%20text.htm
[C8] U.S. National Library of Medicine and the National Institute of Health. Gum Disease. Aug. 2008. 25 Aug. 2008 http://www.nlm.nih.gov/medlineplus/gumdisease.html.
[C9] Pavlica Zlatko. “Periodontal Disease and Microbial Communities.” European Companion Animal Health 2006.
[C10] Saini S, et al. “Microbial Flora in Orodental Infections.” Indian Journal of Medical Microbiology 21:2 (2003) 111-114.
[C11] Ohta H, et al. “Microbial interactions and the development of periodontal disease.” Journal of Periodontal Research 26 (1991) 255-257.
[C12] “Periodontal Disease.” Oral Pathology. Jul. 2008. 25 Aug. 2008 http://www.oralbacteria.com/index.php?option=com_content&task=view&id=25&Itemid=31.
[C13] Lamont Richard J. and Jenkinson Howard F. “Life Below the Gum Line: Pathogenic Mechanisms of Porphyromonas gingivalis.” Microbiology and Molecular Biology Reviews 62:4 (1998) 1244-1263.
[C14] Hirano Yasuyuki, et al. “The cell extract of Porphyromonas gingivalis promotes attachment of Prevotella nigrescens cells to hydroxyapatite.” Journal of Oral Science 45:2 (2003) 99-106.
[C15] Sela Michael N. “Role of Treponema Denticola in Periodontal Diseases.” Critical Reviews in Oral Biology and Mediicne 12:5 (2001) 399-413.
[C16] Vesey Peter M. and Kuramitsu Howard K. “Genetic analysis of Treponema denticola ATCC 35405 biofilm formation.” Microbiology 150 (2004) 2401-2407.
[C17] Los Alamons National Laboratory. “Actinomyces naeslundii MG1.” 2006. 25 Aug. 2008 http://www.oralgen.lanl.gov/oralgen/bacteria/anae/
[C18] Takahashi N. and Yamada T. “Glucose and Lactate Metabolism by Actinomyces Naeslundii.” Critical Reviews in Oral Biology and Medicine 10:4 (1999) 487-503.
[C19] Ge S, Wu YF, Liu TJ, Meng S, Zhao L. “Study of the correlation between moderately and severely chronic periodontitis and coronary heart disease”, Hua Xi Kou Qiang Yi Xue Za Zhi. 2008 Jun;26(3):262-6. PMID: 18705507 http://www.ncbi.nlm.nih.gov/pubmed/18705507?ordinalpos=18&itool=EntrezSystem2.PEntrez.Pubmed.Pubmed_ResultsPanel.Pub
[C20] Chukhlovin AB, Solovyova AM, Matelo SK, Kobiyasova IV, Morosova EB, Hokhlacheva AV, Teplyakov BG, Syssoev KA, Konstantinova VE, Matelo LN, Totolian AA. “Bacterial markers of periodontal diseases and their practical significance in dentistry.” Bull Exp Biol Med. 2007 Oct;144(4):546-50. PMID: 18642710 http://www.ncbi.nlm.nih.gov/pubmed/18642710?ordinalpos=1&itool=EntrezSystem2.PEntrez.Pubmed.Pubmed_ResultsPanel.Pubmed_RVDocSum
[D1] Wilson, M. Bacteriology of Humans: an ecological perspective. Blackwell Publishing LTYD. MA. 2008
[D2] MM Danser, S Mantilla Gomez, GA Van Der Weijden (2003) Tongue coating and tongue brushing: a literature review
[D3] A. Bordas, R. Mcnab, A. Staples, J. Bowman, J. Kanapka, M. Bosma (2008) Impact of different tongue cleaning methods on the bacterial load of the tongue dorsum. Archives of Oral Biology 53, SUPPL. I (2008) S13-S18
[D4] Pennisi, Elizabeth (2005) A Mouthful of Microbes. Science 25 March 2005: Vol.307. no. 5717, pp 1899-1901
[D5] Aas. J.A., Paster, B.J.. Stokes, L.N., Olsen, I. and Dewhirst, F.E. (2005) Defining the normal bacterial flora of the oral cavity. J Clin Micriobiol 43, 5721-32
[D6] Kazor, C.E, Mitchell, P.M. . Lee, A.M. , Stokes, Loesche, W.J., Dewhirst, F.E. and Paster, B.J. (2003) Diversity of bacterial populations on the tongue dorsa of patients with halitosis and healthy patients. J Clin Micriobiol 41, 558-63
[D7] MedlinePlus Medical Encyclopedia: Glossitis. Uppaluri. January 30th 2007. American Accreditation HealthCare Commission. August 20th 2008. < http://www.nlm.nih.gov/medlineplus/ency/article/001053.htm>.
[D8] MedlinePlus Medical Encyclopedia: Herpes Simplex. Zieve, Juhn, Eltz, Stacy. January 30th 2007. American Accreditation HealthCare Commission. August 20th 2008. <http://www.nlm.nih.gov/MEDLINEPLUS/ency/article/001324.htm>
[D9] Rosenberg E. “From Dental Plaque to Oral Biofilm: A Molecular Odyssey.” Microbial Ecology and Infectious Disease. London J. Washington: American Society for Microbiology, 1999. 53-63.
[D10] Bite Sized Tutorials, Streptococci and Oral Streptococci. Newcastle University. August 21st 2008. <http://www.ncl.ac.uk/dental/oralbiol/oralenv/tutorials/streps.htm>
[D11] Method for Inhibiting Yeast Growth Invention. August 21st 2008. <http://www.freshpatents.com/Method-for-inhibiting-yeast-growth-dt20051208ptan20050271640.php>
[D12] Antimicrobial Peptides in the Oral Environment: Expression and Function in Health and Disease. Dale, Fredericks. July 2005. University of Washington. August 21st 2008. <http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1479858>
[D13] Sriprakash KS. “Oral Streptococcal BLIS: Heterogeneity of the Effector Molecules and Potential Role in the Prevention of Streptococcal Infections.” Streptococci: New Insights into an Old Enemy. Tagg J, Wescombe P, Burton J. San Diego: ELSEVIER B.V, 2006. 347-350.
[E1] Hull M.W., Chow A.W. (2007) “Indigenous Microflora and Innate Immunity of the Head and Neck.” Infectious Disease Clinics of North America 21: 265-282.
[E2] Pillsbury H.C., Rose A.S. (2006) “Biology of the Ears, Nose, and Throat: Throat.” The Merck Manual: Online Medical Library (http://www.merck.com/mmhe/sec19/ch217/ch217d.html)
[E3] Pericone C.D., Overweg K., Hermans P.W.M., Weiser J.N. (200). “Inhibitory and bactericidal Effects of Hydrogen Peroxide Production by Streptococcus pneumoniae on Other Inhabitants of the Upper Respiratory Tract.” Infection and Immunity 68: 3990-3997.
[E4] Lysenko E.S., Ratner A.J., Nelson A.J., Weiser J.N. (2005) “The Role of Innate Immune Responses in the Outcome of the Interspecies Competition for Colonization of Mucosal Surfaces.” PLoS Pathogens 1(1): e1 doi:10.1371/journal.ppat.0010001
[E5] Ohsaki Y, Tachibana M, Awaya T, Kuroki M, Itoh Y. (2008). “Recovery of susceptibility to penicillin G (PEN-G) in clinical isolates of Streptococcus pneumoniae despite increased accumulation of pbp gene alterations.” International Journal of Antimicrobial Agents. In Press, Corrected Proof, Available Online August 19, 2008. (http://www.sciencedirect.com/science/article/B6T7H-4T7W3FV-4/2/8c966b9c9574b58ab2a3c1704aa7b2a4)
[E6] Roscoe DL, Chow AW. Normal flora and mucosal immunity of the head and neck. Infect Dis Clin North Am 1988;2(1):1–19.
[E7] Roscoe DL, Chow AW. Normal flora and mucosal immunity of the head and neck. Infect Dis Clin North Am 1988;2(1):6–7
[E8] Greenfield S, Sheehe PR, Feldman HA. Meningococcal carriage in a population of ‘‘normal’’ families. J Infect Dis 1971;123(1):67–73.
[E9] Caugant DA, Hoiby EA, Magnus P, et al. Asymptomatic carriage of Neisseria meningitides in a randomly sampled population. J Clin Microbiol 1994;32(2):323–30.
[E10] Hull, M., Chow, A. “Indigenous Microflora and Innate Immunity of the Head and Neck.” Infect Dis Clin N Am 21. 2007. 265–282
[E11] McCarthy C, Snyder ML, Parker RB. The indigenous oral flora of man. I. The newborn to the 1-year-old infant. Arch Oral Biol 1965;10:61–70.
[E12] Smith DJ, Anderson JM, King WF, et al. Oral streptococcal colonization of infants. Oral Microbiol Immunol 1993;8(1):1–4.
[E13] Hughes CV, Kolenbrander PE, Andersen RN, et al. Coaggregation properties of human oral Veillonella spp.: relationship to colonization site and oral ecology. Appl Environ Microbiol 1988;54(8):1957–63.
[E14] Delwiche EA, Pestka JJ, Tortorello ML. The Veillonellae: gram-negative cocci with a unique physiology. Annu Rev Microbiol 1985;39:175–93.
[E15] Lamont JT. Mucus: the front line of intestinal mucosal defense. AnnNYAcad Sci 1992;664: 190–201.
[E16] Kolenbrander PE, London J. Adhere today, here tomorrow: oral bacterial adherence. J Bacteriol 1993;175(11):3247–52.
[E17] Jenkinson HF, Demuth DR. Structure, function and immunogenicity of streptococcal antigen I/II polypeptides. Mol Microbiol 1997;23(2):183–90.
[E18] Joh D, Wann ER, Kreikemeyer B, et al. Role of fibronectin-binding MSCRAMMs in bacterial adherence and entry into mammalian cells. Matrix Biol 1999;18(3):211–23.
[E19] Medzhitov R, Janeway C Jr. Innate immune recognition: mechanisms and pathways. Immunol Rev 2000;173:89–97.
[E20] Akira S, Hemmi H. Recognition of pathogen-associated molecular patterns by TLR family. Immunol Lett 2003;85(2):85–95.
[E21] Beutler B, Hoebe K, Du X, et al. How we detect microbes and respond to them: the Toll-like receptors and their transducers. J Leukoc Biol 2003;74(4):479–85.
[E22] Kopp E, Medzhitov R. Recognition of microbial infection by Toll-like receptors. Curr Opin Immunol 2003;15(4):396–401.
[E23] Takeda K, Kaisho T, Akira S. Toll-like receptors. Annu Rev Immunol 2003;21:335 76.
[E24] Russell MW, Mestecky J. Induction of the mucosal immune response. Rev Infect Dis 1988; 10(Suppl 2):S440–6.
Edited by Mason Chen, Diana Kirchmann, Alvin Kwong, Lydi Martinez, Mei Ng, Gabriel Tran, Kristen Watanabe, Kathryn Yee students of Rachel Larsen